


Vitor Lopes dos Santos
@vitorlds.bsky.social
Reposted by Vitor Lopes dos Santos

This is beautiful work and quite the discovery. This also plays nicely with the idea that superficial place cells are rigid and deep are flexible. Seems to suggest that the flexibility is more about past rep's integrating with newer rep's, than the opposite.
October 2, 2025 at 5:52 PM
This is beautiful work and quite the discovery. This also plays nicely with the idea that superficial place cells are rigid and deep are flexible. Seems to suggest that the flexibility is more about past rep's integrating with newer rep's, than the opposite.
Thank you Sanja!
June 7, 2025 at 7:20 PM
Thank you Sanja!
The embedding enables a cool experiment: start with a tetrode in CA1 pyr, lower it gradually, and project the signal at each step.
The tetrode follows the predicted path, and the gamma profiles match the silicon probe data beautifully at each step.
See one going from CA1 pyr to DG midmol below!
The tetrode follows the predicted path, and the gamma profiles match the silicon probe data beautifully at each step.
See one going from CA1 pyr to DG midmol below!

June 6, 2025 at 11:38 AM
The embedding enables a cool experiment: start with a tetrode in CA1 pyr, lower it gradually, and project the signal at each step.
The tetrode follows the predicted path, and the gamma profiles match the silicon probe data beautifully at each step.
See one going from CA1 pyr to DG midmol below!
The tetrode follows the predicted path, and the gamma profiles match the silicon probe data beautifully at each step.
See one going from CA1 pyr to DG midmol below!
If you're interested in the code, see: github.com/vitorlds/Hip...
I’ll be updating it soon with a newer Python version and more data!
(13/13)
I’ll be updating it soon with a newer Python version and more data!
(13/13)

GitHub - vitorlds/Hipp-LFP-embedding: Code for Lopes-dos-Santos et al. (2025) Cell Reports
Code for Lopes-dos-Santos et al. (2025) Cell Reports - vitorlds/Hipp-LFP-embedding
github.com
June 5, 2025 at 5:10 PM
If you're interested in the code, see: github.com/vitorlds/Hip...
I’ll be updating it soon with a newer Python version and more data!
(13/13)
I’ll be updating it soon with a newer Python version and more data!
(13/13)
Overall, I had a lot of fun preparing this manuscript.
It started as a side project to support various experiments in the lab, but it grew into its own thing.
Huge thanks to Demi and David for working with me on this at @mrcbndu.ox.ac.uk, and to our reviewers for helping improve the paper.
(12/13)
It started as a side project to support various experiments in the lab, but it grew into its own thing.
Huge thanks to Demi and David for working with me on this at @mrcbndu.ox.ac.uk, and to our reviewers for helping improve the paper.
(12/13)
June 5, 2025 at 5:10 PM
Overall, I had a lot of fun preparing this manuscript.
It started as a side project to support various experiments in the lab, but it grew into its own thing.
Huge thanks to Demi and David for working with me on this at @mrcbndu.ox.ac.uk, and to our reviewers for helping improve the paper.
(12/13)
It started as a side project to support various experiments in the lab, but it grew into its own thing.
Huge thanks to Demi and David for working with me on this at @mrcbndu.ox.ac.uk, and to our reviewers for helping improve the paper.
(12/13)
In short: LFPs carry rich circuit information.
We mapped oscillations and neuronal firing across CA1–DG layers.
This opens new doors for identifying oscillations, circuit motifs, and homogenize layer identification across datasets.
Check out the paper: doi.org/10.1016/j.ce...
(11/13)
We mapped oscillations and neuronal firing across CA1–DG layers.
This opens new doors for identifying oscillations, circuit motifs, and homogenize layer identification across datasets.
Check out the paper: doi.org/10.1016/j.ce...
(11/13)
Redirecting
doi.org
June 5, 2025 at 5:10 PM
In short: LFPs carry rich circuit information.
We mapped oscillations and neuronal firing across CA1–DG layers.
This opens new doors for identifying oscillations, circuit motifs, and homogenize layer identification across datasets.
Check out the paper: doi.org/10.1016/j.ce...
(11/13)
We mapped oscillations and neuronal firing across CA1–DG layers.
This opens new doors for identifying oscillations, circuit motifs, and homogenize layer identification across datasets.
Check out the paper: doi.org/10.1016/j.ce...
(11/13)
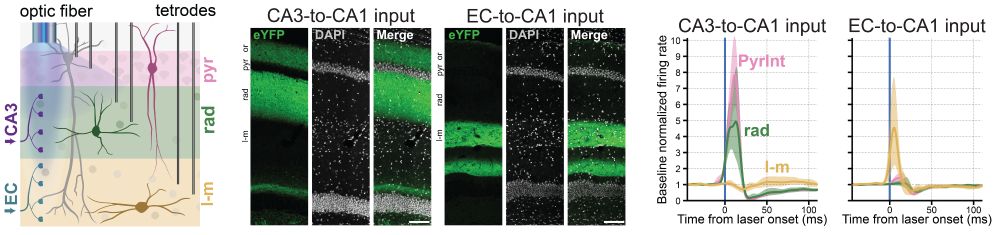
Using optogenetics, we tested how CA1 interneurons across layers respond to upstream input.
CA3 terminal stimulation strongly activated pyramidale (PyrInt) and radiatum (rad) interneurons, while EC terminal stimulation strongly drove LM interneurons.
(10/13)
CA3 terminal stimulation strongly activated pyramidale (PyrInt) and radiatum (rad) interneurons, while EC terminal stimulation strongly drove LM interneurons.
(10/13)
June 5, 2025 at 5:10 PM
Using optogenetics, we tested how CA1 interneurons across layers respond to upstream input.
CA3 terminal stimulation strongly activated pyramidale (PyrInt) and radiatum (rad) interneurons, while EC terminal stimulation strongly drove LM interneurons.
(10/13)
CA3 terminal stimulation strongly activated pyramidale (PyrInt) and radiatum (rad) interneurons, while EC terminal stimulation strongly drove LM interneurons.
(10/13)
Using embedding-guided tetrodes, Demi recorded interneurons in sparse CA1 layers: rad and LM (btw great dataset, stay tuned for future work on this!).
Firing behavior was layer-specific; e.g., LM interneurons were barely active in ripples.
Preferred theta phase also varied across layers.
(9/13)
Firing behavior was layer-specific; e.g., LM interneurons were barely active in ripples.
Preferred theta phase also varied across layers.
(9/13)

June 5, 2025 at 5:10 PM
Using embedding-guided tetrodes, Demi recorded interneurons in sparse CA1 layers: rad and LM (btw great dataset, stay tuned for future work on this!).
Firing behavior was layer-specific; e.g., LM interneurons were barely active in ripples.
Preferred theta phase also varied across layers.
(9/13)
Firing behavior was layer-specific; e.g., LM interneurons were barely active in ripples.
Preferred theta phase also varied across layers.
(9/13)
I love this figure
By projecting tetrode LFPs from several mice onto the embedding (estimating each tetrode’s “depth”), we recover gradually changing theta and ripple waveforms, as if recorded with a single linear probe.
No need for alignment or histology. Just the signals and the embedding.
(8/13)
By projecting tetrode LFPs from several mice onto the embedding (estimating each tetrode’s “depth”), we recover gradually changing theta and ripple waveforms, as if recorded with a single linear probe.
No need for alignment or histology. Just the signals and the embedding.
(8/13)

June 5, 2025 at 5:10 PM
I love this figure
By projecting tetrode LFPs from several mice onto the embedding (estimating each tetrode’s “depth”), we recover gradually changing theta and ripple waveforms, as if recorded with a single linear probe.
No need for alignment or histology. Just the signals and the embedding.
(8/13)
By projecting tetrode LFPs from several mice onto the embedding (estimating each tetrode’s “depth”), we recover gradually changing theta and ripple waveforms, as if recorded with a single linear probe.
No need for alignment or histology. Just the signals and the embedding.
(8/13)
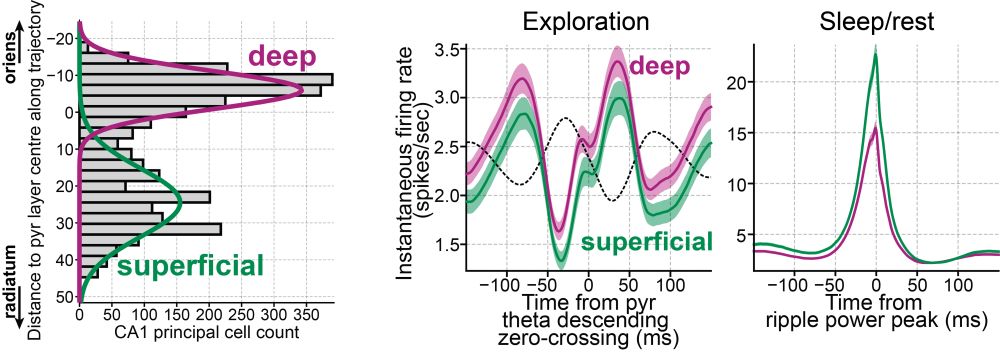
Zooming in, the embedding also separates deep vs superficial CA1 pyramidal cells.
These groups differ in firing rates (awake and sleep), theta modulation, and coupling to gamma and ripple.
This let us replicate findings from fancy silicon probes using tetrodes, and extend them further.
(7/13)
These groups differ in firing rates (awake and sleep), theta modulation, and coupling to gamma and ripple.
This let us replicate findings from fancy silicon probes using tetrodes, and extend them further.
(7/13)
June 5, 2025 at 5:10 PM
Zooming in, the embedding also separates deep vs superficial CA1 pyramidal cells.
These groups differ in firing rates (awake and sleep), theta modulation, and coupling to gamma and ripple.
This let us replicate findings from fancy silicon probes using tetrodes, and extend them further.
(7/13)
These groups differ in firing rates (awake and sleep), theta modulation, and coupling to gamma and ripple.
This let us replicate findings from fancy silicon probes using tetrodes, and extend them further.
(7/13)