



Timothy Saunders
@timesaunders.bsky.social
Professor at Warwick Medical School interested in how complex shape emerges during embryo development. Will work on anything that can be imaged live :)
Of course, biology likes to be complicated. The above work generally takes the microtubule interaction between nuclei to be repulsive - but this doesn’t have to be the case (www.nature.com/articles/s41...). Future work will hopefully reveal how general these principles are.
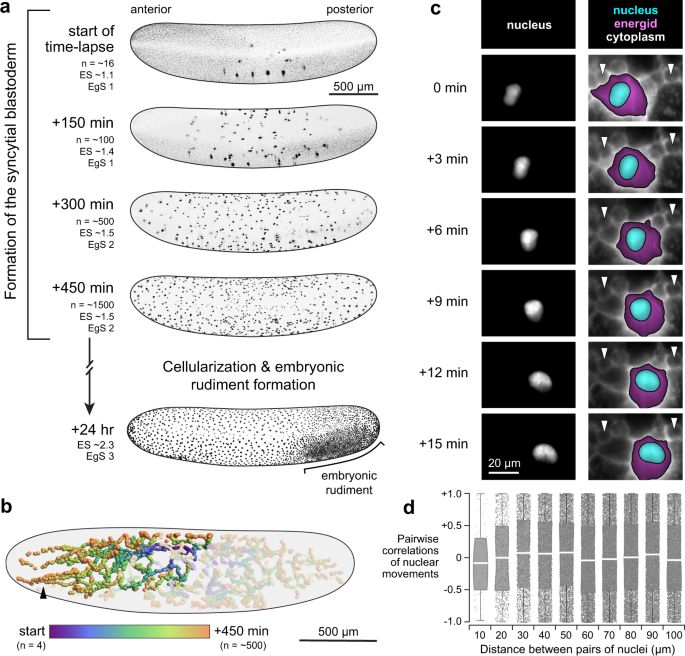
Nuclear speed and cycle length co-vary with local density during syncytial blastoderm formation in a cricket - Nature Communications
Early in insect embryo development, many nuclei share one large cell, travel varied paths and self-organize into a single layer. Donoughe et al. illuminate this process with live-imaging, modeling, an...
www.nature.com
March 13, 2025 at 11:07 AM
Of course, biology likes to be complicated. The above work generally takes the microtubule interaction between nuclei to be repulsive - but this doesn’t have to be the case (www.nature.com/articles/s41...). Future work will hopefully reveal how general these principles are.
With Ivo Telley, we have looked at packing of nuclei in ex vivo systems (journals.biologists.com/dev/article/..., elifesciences.org/articles/90541). It’s exciting to see how quantitative measurements reveal deeper insight into how biological systems reliably generate complex form from simple rules.
Aster repulsion drives short-ranged ordering in the Drosophila syncytial blastoderm
Summary: The multinucleated embryo of the fruit fly D. melanogaster distributes nuclei with the help of centrosome-organised microtubule asters, with ordering dynamically adapting to synchronous nucle...
journals.biologists.com
March 13, 2025 at 11:05 AM
With Ivo Telley, we have looked at packing of nuclei in ex vivo systems (journals.biologists.com/dev/article/..., elifesciences.org/articles/90541). It’s exciting to see how quantitative measurements reveal deeper insight into how biological systems reliably generate complex form from simple rules.
Such topological behaviour has also been shown to potentially play a role in cell extrusion (www.nature.com/articles/nat...). I wrote a short commentary on this, which is hopefully accessible for the more general audience (www.nature.com/articles/s41...).
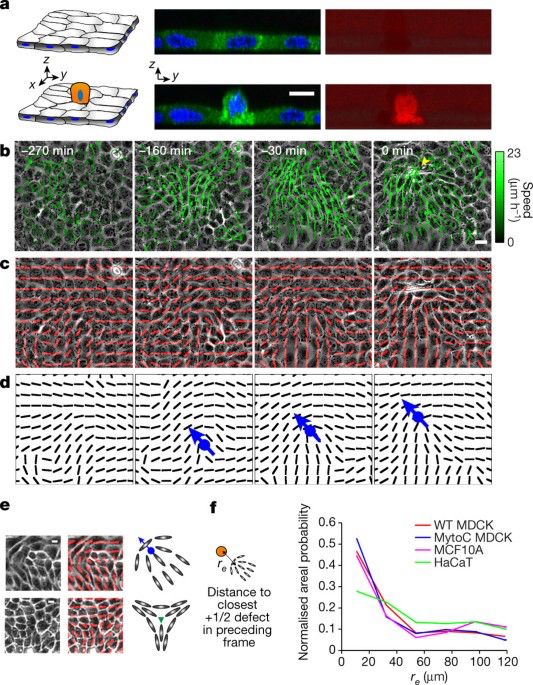
Topological defects in epithelia govern cell death and extrusion - Nature
By modelling epithelial cells as active nematic liquid crystals, stresses induced at the sites of topological defects are found to be the primary drivers of extrusion and cell death.
www.nature.com
March 13, 2025 at 11:03 AM
Such topological behaviour has also been shown to potentially play a role in cell extrusion (www.nature.com/articles/nat...). I wrote a short commentary on this, which is hopefully accessible for the more general audience (www.nature.com/articles/s41...).
A wonderful day. Thanks so much for the invitation
December 13, 2024 at 8:23 PM
A wonderful day. Thanks so much for the invitation
Hi Joachim. Can you add me - we do optogenetics ( elifesciences.org/articles/26258), laser ablation (www.cell.com/current-biol... and any quantitative tools we can use in fish and flies.

Decoding temporal interpretation of the morphogen Bicoid in the early Drosophila embryo
Cell fates endowed by higher Bicoid concentration require input for longer duration, demonstrating a temporally non-linear morphogen-mediated pattern formation.
elifesciences.org
November 20, 2024 at 10:37 AM
Hi Joachim. Can you add me - we do optogenetics ( elifesciences.org/articles/26258), laser ablation (www.cell.com/current-biol... and any quantitative tools we can use in fish and flies.
Just discovering how clunky Bluesky is for this :(
November 17, 2024 at 8:40 PM
Just discovering how clunky Bluesky is for this :(
Our collaborators - the Weil lab - primarily focused on the first question. They revealed that localisation of bicoid mRNA localisation to P bodies inhibits translation. Chemically releasing bicoid from the P bodies resulted in translation in the oocyte…
November 17, 2024 at 8:40 PM
Our collaborators - the Weil lab - primarily focused on the first question. They revealed that localisation of bicoid mRNA localisation to P bodies inhibits translation. Chemically releasing bicoid from the P bodies resulted in translation in the oocyte…
We generated a Bicoid::Suntag reporter line. Importantly, this could address two important questions: (i) is bicoid translated in the oocyte, and if not, what is regulating this process?; (ii) where and when is bicoid translated in the embryo? (20)
November 17, 2024 at 8:40 PM
We generated a Bicoid::Suntag reporter line. Importantly, this could address two important questions: (i) is bicoid translated in the oocyte, and if not, what is regulating this process?; (ii) where and when is bicoid translated in the embryo? (20)
We were motivated by the development of SunTag (PMID: 25307933), especially its application to the living Drosophila embryo by the Mounia lab (PMID: 33927056). (19)
November 17, 2024 at 8:40 PM
We were motivated by the development of SunTag (PMID: 25307933), especially its application to the living Drosophila embryo by the Mounia lab (PMID: 33927056). (19)
It has remained an open question as to precisely when and where bicoid mRNA is translated. If bicoid mRNA is not translated in the oocyte, what is the mechanism? This motivated us to revisit the “S” part of the SDD model. (18/22)
November 17, 2024 at 8:40 PM
It has remained an open question as to precisely when and where bicoid mRNA is translated. If bicoid mRNA is not translated in the oocyte, what is the mechanism? This motivated us to revisit the “S” part of the SDD model. (18/22)
These direct measures of motion reveal that the “diffusive” part of the SDD model needs adaptation (e.g., spatial and temporal change), but Bicoid does move through the embryo sufficiently fast to generate a gradient. (18/22)
November 17, 2024 at 8:40 PM
These direct measures of motion reveal that the “diffusive” part of the SDD model needs adaptation (e.g., spatial and temporal change), but Bicoid does move through the embryo sufficiently fast to generate a gradient. (18/22)
With Thorsten Wohland, we used FCS at multiple locations and time points on Bicoid. We demonstrated different rates of diffusivity in the anterior and posterior regions (PMID: 38345326). (17/22)
November 17, 2024 at 8:40 PM
With Thorsten Wohland, we used FCS at multiple locations and time points on Bicoid. We demonstrated different rates of diffusivity in the anterior and posterior regions (PMID: 38345326). (17/22)
However, these measurements were essentially averaging over the whole system – what is happening with Bicoid at more local (e.g., within a nuclear domain) scales? (16/22)
November 17, 2024 at 8:40 PM
However, these measurements were essentially averaging over the whole system – what is happening with Bicoid at more local (e.g., within a nuclear domain) scales? (16/22)
While all the models proposed can generate an exponentially-decaying profile, they all made different predictions for how old Bicoid would be at different locations. Only the SDD model was consistent with the overall behaviour of the protein age. (15/22)
November 17, 2024 at 8:40 PM
While all the models proposed can generate an exponentially-decaying profile, they all made different predictions for how old Bicoid would be at different locations. Only the SDD model was consistent with the overall behaviour of the protein age. (15/22)
During my post-doc and into my own lab we developed a Bicoid-timer reporter. By tagging Bicoid with two fluorescent proteins with different folding rates, we could effectively measure the age (i.e., ratio of the two fluorophore signals) of Bicoid at different locations (PMID: 30181144). (14/22)
November 17, 2024 at 8:40 PM
During my post-doc and into my own lab we developed a Bicoid-timer reporter. By tagging Bicoid with two fluorescent proteins with different folding rates, we could effectively measure the age (i.e., ratio of the two fluorophore signals) of Bicoid at different locations (PMID: 30181144). (14/22)
Fluorescence correlation spectroscopy (FCS) of Bicoid::eGFP revealed that the dynamics were more complicated than inferred from FRAP. In particular, an effective diffusion coefficient of around 7um2s-1 (PMID: 20712981). This measurement was more in line with expectations from the SDD model. (13/22)
November 17, 2024 at 8:40 PM
Fluorescence correlation spectroscopy (FCS) of Bicoid::eGFP revealed that the dynamics were more complicated than inferred from FRAP. In particular, an effective diffusion coefficient of around 7um2s-1 (PMID: 20712981). This measurement was more in line with expectations from the SDD model. (13/22)