



Matthew Taliaferro
@jmtali.bsky.social
RNA biologist at University of Colorado Anschutz Medical Campus http://www.taliaferrolab.com
How does one review his paper if he’s infallible?

May 9, 2025 at 12:20 AM
How does one review his paper if he’s infallible?
If we replace wildtype TDP-43 with an ALS mutant TDP-43, we again see that the same RNAs become aberrantly neurite-enriched. So this could be happening in ALS patient cells, but whether it makes contributions to patient phenotypes is another question for another day.

March 3, 2025 at 4:00 PM
If we replace wildtype TDP-43 with an ALS mutant TDP-43, we again see that the same RNAs become aberrantly neurite-enriched. So this could be happening in ALS patient cells, but whether it makes contributions to patient phenotypes is another question for another day.
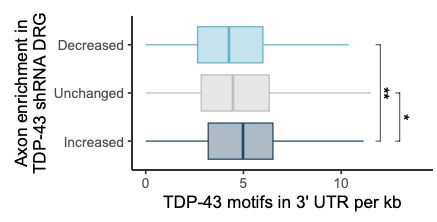
Does any of this have anything to do with ALS? Maybe. We see the same RNA localization defects happening in primary mouse motor neurons and human iPS-derived motor neurons upon TDP-43 loss.
March 3, 2025 at 4:00 PM
Does any of this have anything to do with ALS? Maybe. We see the same RNA localization defects happening in primary mouse motor neurons and human iPS-derived motor neurons upon TDP-43 loss.
260mers that drove TDP-43-dependent localization and were bound by TDP-43 in vitro also showed TDP-43-dependent changes in stability, directly linking all three processes. All of these qualities were eliminated if TDP-43 motifs in the 260mers were eliminated.



March 3, 2025 at 3:59 PM
260mers that drove TDP-43-dependent localization and were bound by TDP-43 in vitro also showed TDP-43-dependent changes in stability, directly linking all three processes. All of these qualities were eliminated if TDP-43 motifs in the 260mers were eliminated.
OK, so *how* is TDP-43 doing this? TDP-43 negatively regulates RNA stability and recent work has linked RNA localization and stability. So is this what's going on? To test this we measured the stability of the same ~10k reporters with and without TDP-43 using SLAM-seq.

March 3, 2025 at 3:59 PM
OK, so *how* is TDP-43 doing this? TDP-43 negatively regulates RNA stability and recent work has linked RNA localization and stability. So is this what's going on? To test this we measured the stability of the same ~10k reporters with and without TDP-43 using SLAM-seq.
Sure enough, 260mers that have TDP-43-dependent localization activity are directly bound by TDP-43, and mutation of TDP-43 motifs within them abrogates both binding and activity.



March 3, 2025 at 3:59 PM
Sure enough, 260mers that have TDP-43-dependent localization activity are directly bound by TDP-43, and mutation of TDP-43 motifs within them abrogates both binding and activity.
OK so we have 260mers that have TDP-43-dependent RNA localization activity. But are they actually bound by TDP-43? To test this, we measured the TDP-43 affinity for the exact same ~10k 260mer RNAs in vitro, allowing us to directly relate binding to activity.

March 3, 2025 at 3:59 PM
OK so we have 260mers that have TDP-43-dependent RNA localization activity. But are they actually bound by TDP-43? To test this, we measured the TDP-43 affinity for the exact same ~10k 260mer RNAs in vitro, allowing us to directly relate binding to activity.
So what distinguishes these bound/active motifs from unbound/inactive motifs? Secondary structure. Bound motifs are significantly more single-stranded. We can predict whether or not a TDP-43 motif regulates RNA localization simply by looking at its secondary structure character.


March 3, 2025 at 3:58 PM
So what distinguishes these bound/active motifs from unbound/inactive motifs? Secondary structure. Bound motifs are significantly more single-stranded. We can predict whether or not a TDP-43 motif regulates RNA localization simply by looking at its secondary structure character.
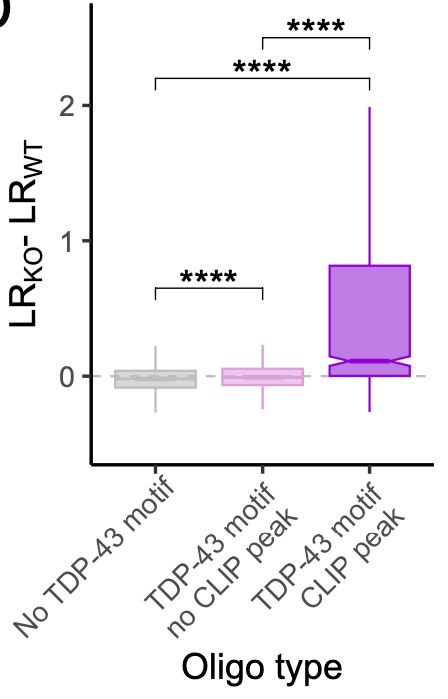
Only some TDP-43 motifs are actually occupied by TDP-43 in cells, and only those motifs made RNAs TDP-43-sensitive. Further, mutation of those motifs abolished activity, while mutation of other non-occupied motifs had no effect.

March 3, 2025 at 3:58 PM
Only some TDP-43 motifs are actually occupied by TDP-43 in cells, and only those motifs made RNAs TDP-43-sensitive. Further, mutation of those motifs abolished activity, while mutation of other non-occupied motifs had no effect.
We found specific pieces of UTRs that were sufficient to make the localization of reporters sensitive to TDP-43. Interestingly, these often overlapped perfectly with known TDP-43 CLIP sites. Their activity was also abolished by mutated TDP-43 motifs within them.

March 3, 2025 at 3:58 PM
We found specific pieces of UTRs that were sufficient to make the localization of reporters sensitive to TDP-43. Interestingly, these often overlapped perfectly with known TDP-43 CLIP sites. Their activity was also abolished by mutated TDP-43 motifs within them.
To answer that, we used an MPRA where we chopped up those UTRs into ~10k 260mer RNA sequences. We then incorporated each of those into a reporter and tested the localization of those reporters with and without TDP-43.

March 3, 2025 at 3:57 PM
To answer that, we used an MPRA where we chopped up those UTRs into ~10k 260mer RNA sequences. We then incorporated each of those into a reporter and tested the localization of those reporters with and without TDP-43.
Further, the 3' UTRs of the 3 exemplar RNAs were sufficient to make the localization of a reporter RNA similarly dependent upon TDP-43. But what about these UTRs wast10
making them TDP-43-sensitive?
making them TDP-43-sensitive?

March 3, 2025 at 3:57 PM
Further, the 3' UTRs of the 3 exemplar RNAs were sufficient to make the localization of a reporter RNA similarly dependent upon TDP-43. But what about these UTRs wast10
making them TDP-43-sensitive?
making them TDP-43-sensitive?
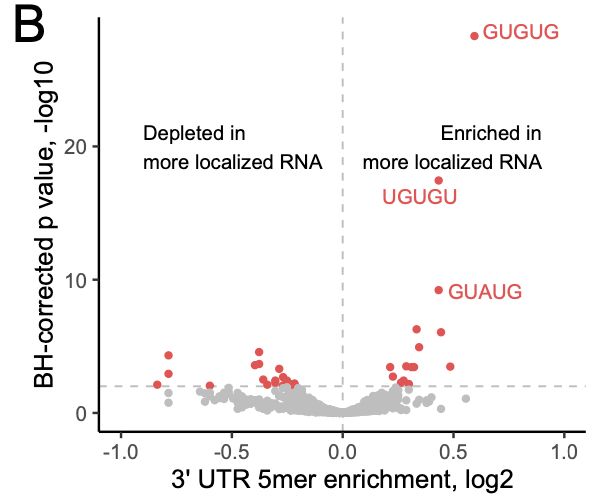
TDP-43 binds GU-rich kmers. 3' UTRs of RNAs that became more neurite-enriched upon TDP-43 loss were very strongly enriched in these motifs as well as CLIP-defined TDP-43 binding sites, suggesting that they are being directly regulated by TDP-43.

March 3, 2025 at 3:57 PM
TDP-43 binds GU-rich kmers. 3' UTRs of RNAs that became more neurite-enriched upon TDP-43 loss were very strongly enriched in these motifs as well as CLIP-defined TDP-43 binding sites, suggesting that they are being directly regulated by TDP-43.
Interestingly, many RNAs became *more* neurite-enriched without TDP-43. This is exemplified by 3 RNAs, Diras1, Ksr2, and Wasf3. Although the total cellular level of these RNAs stays ~ the same, in neurites they go up 20-fold without TDP-43.


March 3, 2025 at 3:57 PM
Interestingly, many RNAs became *more* neurite-enriched without TDP-43. This is exemplified by 3 RNAs, Diras1, Ksr2, and Wasf3. Although the total cellular level of these RNAs stays ~ the same, in neurites they go up 20-fold without TDP-43.
Interestingly, many RNAs became *more* neurite-enriched without TDP-43. This is exemplified by 3 RNAs, Diras1, Ksr2, and Wasf3. Although the total cellular level of these RNAs stays ~ the same, in neurites they go up 20-fold without TDP-43.


March 3, 2025 at 3:56 PM
Interestingly, many RNAs became *more* neurite-enriched without TDP-43. This is exemplified by 3 RNAs, Diras1, Ksr2, and Wasf3. Although the total cellular level of these RNAs stays ~ the same, in neurites they go up 20-fold without TDP-43.
To ask how TDP-43 regulates RNA localization, we then analyzed neurite enrichments changed with and without dox-inducible loss of cellular TDP-43.

March 3, 2025 at 3:56 PM
To ask how TDP-43 regulates RNA localization, we then analyzed neurite enrichments changed with and without dox-inducible loss of cellular TDP-43.
We used a compartmentalized culture system to separate cell bodies and neurites. By profiling the RNA content of each fraction using RNAseq, we learn the relative abundance of each RNA in neurites vs. soma.

March 3, 2025 at 3:56 PM
We used a compartmentalized culture system to separate cell bodies and neurites. By profiling the RNA content of each fraction using RNAseq, we learn the relative abundance of each RNA in neurites vs. soma.
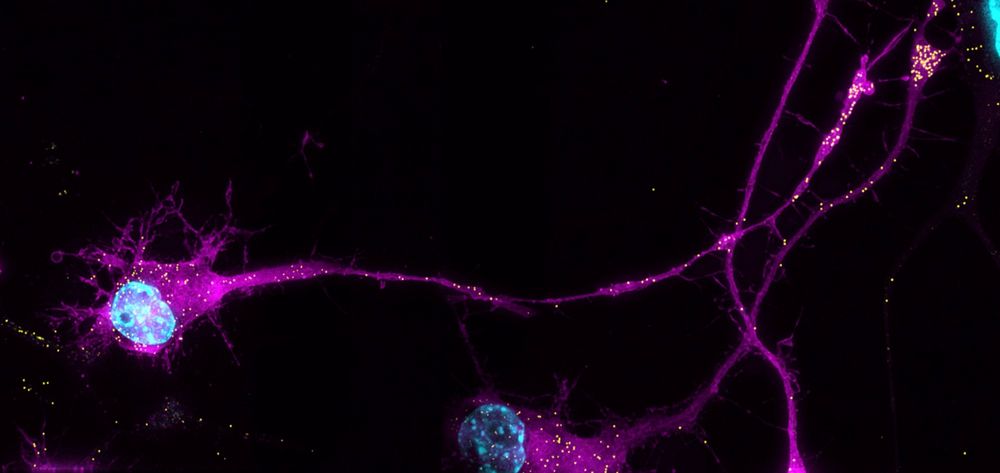
Excited to share our latest work! We asked if and how TDP-43 regulates RNA transport in neurons and if this is misregulated in ALS. We found that it works to keep RNAs *out* of neurites, and that loss of TDP-43 activity results in specific RNAs aberrantly accumulating there.
March 3, 2025 at 3:55 PM
Excited to share our latest work! We asked if and how TDP-43 regulates RNA transport in neurons and if this is misregulated in ALS. We found that it works to keep RNAs *out* of neurites, and that loss of TDP-43 activity results in specific RNAs aberrantly accumulating there.
Save the date for the next edition of this wonderful meeting. We look forward to seeing you in Portugal!!

February 28, 2025 at 3:53 PM
Save the date for the next edition of this wonderful meeting. We look forward to seeing you in Portugal!!
From this we can conclude that I am:
A. extremely arrogant about my own writing
B. an idiot
C. both
A. extremely arrogant about my own writing
B. an idiot
C. both

February 1, 2025 at 5:33 PM
From this we can conclude that I am:
A. extremely arrogant about my own writing
B. an idiot
C. both
A. extremely arrogant about my own writing
B. an idiot
C. both
Come be our colleague! @cu-bmg.bsky.social is running an open rank search for a structural biologist specializing in cryo-EM/ET. This is a great opportunity in a highly collegial and collaborative department.
Apply here: cu.taleo.net/careersectio...
Apply here: cu.taleo.net/careersectio...

January 27, 2025 at 4:36 PM
Come be our colleague! @cu-bmg.bsky.social is running an open rank search for a structural biologist specializing in cryo-EM/ET. This is a great opportunity in a highly collegial and collaborative department.
Apply here: cu.taleo.net/careersectio...
Apply here: cu.taleo.net/careersectio...

One of my favorite things about college football is that this may be enough to get him fired. The sheer irrationality of it all.

November 30, 2024 at 10:26 PM
One of my favorite things about college football is that this may be enough to get him fired. The sheer irrationality of it all.
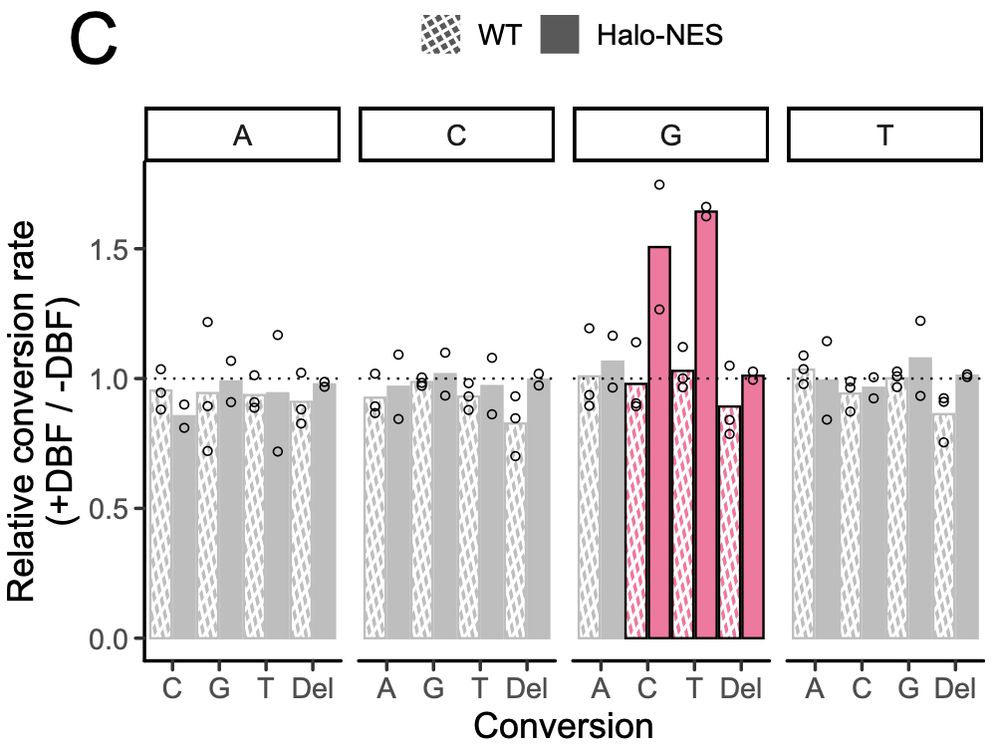
Excitingly, we detect G conversions in the RNA from these embryos, and these conversions also show spatial specificity as MALAT1 had more conversions with a nuclear Halo fusion and GAPDH had more conversions with a cytoplasmic Halo fusion.
[fin] <-- (zebrafish joke)
[fin] <-- (zebrafish joke)

November 13, 2024 at 4:52 PM
Excitingly, we detect G conversions in the RNA from these embryos, and these conversions also show spatial specificity as MALAT1 had more conversions with a nuclear Halo fusion and GAPDH had more conversions with a cytoplasmic Halo fusion.
[fin] <-- (zebrafish joke)
[fin] <-- (zebrafish joke)
Finally, we asked if we could push OINC-seq where no other RNA proximity method can go: inside live animals! We made zebrafish lines expressing Halo fusion proteins and performed the labeling in live embryos.

November 13, 2024 at 4:51 PM
Finally, we asked if we could push OINC-seq where no other RNA proximity method can go: inside live animals! We made zebrafish lines expressing Halo fusion proteins and performed the labeling in live embryos.