




Elsa Abs
@elsa-abs.bsky.social
Researcher CNRS @LSCE_IPSL - Earth scientist and microbial ecologist - @ERC_Research for project GAMEchange - they/iel 🇫🇷 🇱🇧 🏳️🌈
https://www.elsaabs.com/
https://www.elsaabs.com/
Wondering where to submit your abstract for #AGU25 @agu.org?
Check out our session co-organized with @kristenobacter.bsky.social (UMass), Ulas Karaoz and Nicola Falco (Berkeley Lab).
In-person invited speakers: @andreasrichter.bsky.social (U of Vienna) & Amilcare Porporato (Princeton) 🤩
Check out our session co-organized with @kristenobacter.bsky.social (UMass), Ulas Karaoz and Nicola Falco (Berkeley Lab).
In-person invited speakers: @andreasrichter.bsky.social (U of Vienna) & Amilcare Porporato (Princeton) 🤩

July 16, 2025 at 8:33 AM
Wondering where to submit your abstract for #AGU25 @agu.org?
Check out our session co-organized with @kristenobacter.bsky.social (UMass), Ulas Karaoz and Nicola Falco (Berkeley Lab).
In-person invited speakers: @andreasrichter.bsky.social (U of Vienna) & Amilcare Porporato (Princeton) 🤩
Check out our session co-organized with @kristenobacter.bsky.social (UMass), Ulas Karaoz and Nicola Falco (Berkeley Lab).
In-person invited speakers: @andreasrichter.bsky.social (U of Vienna) & Amilcare Porporato (Princeton) 🤩
5. Global heterogeneity of the eco-evolutionary effect
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)

June 20, 2025 at 4:35 PM
5. Global heterogeneity of the eco-evolutionary effect
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
4. Implication for global soil C projections
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)

June 20, 2025 at 4:35 PM
4. Implication for global soil C projections
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
2. Evolutionary result
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)

June 20, 2025 at 4:35 PM
2. Evolutionary result
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
1. The eco-evolutionary model
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)

June 20, 2025 at 4:35 PM
1. The eco-evolutionary model
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
New paper out in Global Change Biology 🤩
doi.org/10.1111/gcb....
We built the first soil carbon model that includes microbial eco-evolution using game theory — and found that adaptation could nearly double global soil carbon loss by 2100. Here is how👇(1/7)
doi.org/10.1111/gcb....
We built the first soil carbon model that includes microbial eco-evolution using game theory — and found that adaptation could nearly double global soil carbon loss by 2100. Here is how👇(1/7)

June 20, 2025 at 4:35 PM
New paper out in Global Change Biology 🤩
doi.org/10.1111/gcb....
We built the first soil carbon model that includes microbial eco-evolution using game theory — and found that adaptation could nearly double global soil carbon loss by 2100. Here is how👇(1/7)
doi.org/10.1111/gcb....
We built the first soil carbon model that includes microbial eco-evolution using game theory — and found that adaptation could nearly double global soil carbon loss by 2100. Here is how👇(1/7)
5. Global heterogeneity of the eco-evolutionary effect
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
June 20, 2025 at 4:27 PM
5. Global heterogeneity of the eco-evolutionary effect
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
We wondered if we could replace eco-evolution with a constant correction. Answer: no. Its effect is uneven—negligible in warm regions, but up to 2× more soil C loss in cold ones. Why? Optimal enzyme allocation responds nonlinearly. (6/7)
4. Implication for global soil C projections
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
June 20, 2025 at 4:27 PM
4. Implication for global soil C projections
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
We thought that microbial adaptation would buffer warming-induced soil C loss. But because it amplifies enzyme production (as shown above), we found that adaptation aggravates the loss—by x1.8 globally. (5/7)
2. Evolutionary result
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
June 20, 2025 at 4:27 PM
2. Evolutionary result
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
We found that in hostile environments (e.g. high mortality, slow uptake), selection favors direct investment in biomass over the riskier strategy of enzyme production. Since warming mainly increases uptake rate, we predicted it would favor stronger enzyme producers. (3/7)
1. The eco-evolutionary model
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
June 20, 2025 at 4:27 PM
1. The eco-evolutionary model
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
We added a trade-off to a classic microbe-soil C model and used adaptive dynamics to evolve enzyme allocation. To prevent freeloaders from taking over, we included implicit spatial structure. Bonus: enzyme production emerges—it’s no longer a free parameter. (2/7)
Gorgeous place to think about eco-evolution. Thanks @lemontree-uofr.bsky.social for the invite!

April 14, 2025 at 9:22 AM
Gorgeous place to think about eco-evolution. Thanks @lemontree-uofr.bsky.social for the invite!
📣 Our special feature highlighting cutting-edge microbial eco-evolutionary research is now live and open for submissions! online.ucpress.edu/elementa/pag.... Submission deadline: September 20, 2025 (but accepted papers will be published on a rolling basis starting immediately).

April 1, 2025 at 1:56 PM
📣 Our special feature highlighting cutting-edge microbial eco-evolutionary research is now live and open for submissions! online.ucpress.edu/elementa/pag.... Submission deadline: September 20, 2025 (but accepted papers will be published on a rolling basis starting immediately).
If you loved (or missed!) our AGU24 session, don’t miss our
#EGU25 session, "Advancing Soil Carbon Predictions"! 🥁 Invited speaker: Elena Shevliakova (Princeton), a leader in integrating microbes into land surface models. Submit abstracts by Jan 15 🗓️: meetingorganizer.copernicus.org/EGU25/sessio...
#EGU25 session, "Advancing Soil Carbon Predictions"! 🥁 Invited speaker: Elena Shevliakova (Princeton), a leader in integrating microbes into land surface models. Submit abstracts by Jan 15 🗓️: meetingorganizer.copernicus.org/EGU25/sessio...

December 17, 2024 at 1:02 AM
If you loved (or missed!) our AGU24 session, don’t miss our
#EGU25 session, "Advancing Soil Carbon Predictions"! 🥁 Invited speaker: Elena Shevliakova (Princeton), a leader in integrating microbes into land surface models. Submit abstracts by Jan 15 🗓️: meetingorganizer.copernicus.org/EGU25/sessio...
#EGU25 session, "Advancing Soil Carbon Predictions"! 🥁 Invited speaker: Elena Shevliakova (Princeton), a leader in integrating microbes into land surface models. Submit abstracts by Jan 15 🗓️: meetingorganizer.copernicus.org/EGU25/sessio...
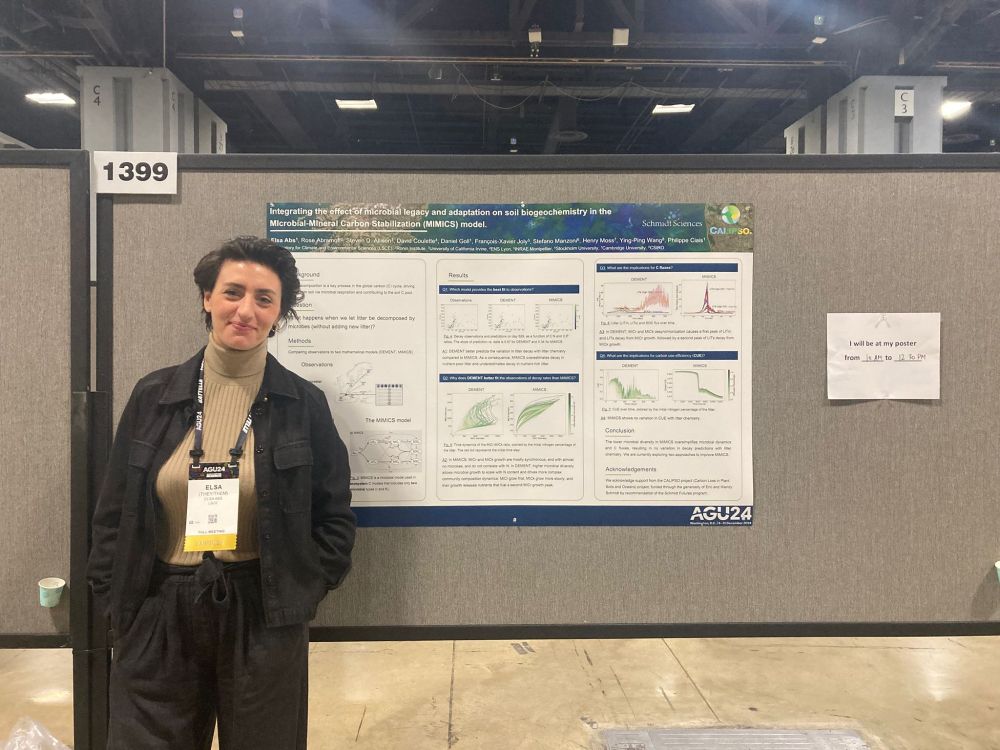
Poster’s up!
Come by Hall B 1399 to chat about integrating microbial diversity and evolution into soil carbon ecosystem models. I’ll be there from 10 AM to 12:30 PM—hope to see you! #AGU24
Come by Hall B 1399 to chat about integrating microbial diversity and evolution into soil carbon ecosystem models. I’ll be there from 10 AM to 12:30 PM—hope to see you! #AGU24
December 11, 2024 at 1:57 PM
Poster’s up!
Come by Hall B 1399 to chat about integrating microbial diversity and evolution into soil carbon ecosystem models. I’ll be there from 10 AM to 12:30 PM—hope to see you! #AGU24
Come by Hall B 1399 to chat about integrating microbial diversity and evolution into soil carbon ecosystem models. I’ll be there from 10 AM to 12:30 PM—hope to see you! #AGU24
I’m at #AGU24 all week! 📋Come to my poster on Wed morning to chat about integrating microbial diversity and adaptation into ecosystem models. 🗣️Come to our 2 oral sessions Wed afternoon to talk about improving soil carbon predictions. ☕️And message me to chat over coffee!
December 9, 2024 at 5:51 PM
I’m at #AGU24 all week! 📋Come to my poster on Wed morning to chat about integrating microbial diversity and adaptation into ecosystem models. 🗣️Come to our 2 oral sessions Wed afternoon to talk about improving soil carbon predictions. ☕️And message me to chat over coffee!
If you signed up for the Soil Carbon Research starter pack and are heading to #AGU2024, don’t miss our session on integrating microbial complexity to advance soil carbon predictions! 🦠🌍 Featuring invited speakers: Christina Kaiser, Jackie Weissman, Katie Rocci, and Will Wieder 🤩.

December 6, 2024 at 6:28 PM
If you signed up for the Soil Carbon Research starter pack and are heading to #AGU2024, don’t miss our session on integrating microbial complexity to advance soil carbon predictions! 🦠🌍 Featuring invited speakers: Christina Kaiser, Jackie Weissman, Katie Rocci, and Will Wieder 🤩.
I was honored to present the IPCC report and France’s specific climate situation today as part of the “Training in the Ecological Transition” for state executives at the Ministry of the Interior, alongside @pfriedling.bsky.social. Hope it inspires meaningful political action!

December 4, 2024 at 5:53 PM
I was honored to present the IPCC report and France’s specific climate situation today as part of the “Training in the Ecological Transition” for state executives at the Ministry of the Interior, alongside @pfriedling.bsky.social. Hope it inspires meaningful political action!
📢 Add to your #AGU2024 schedule our 3 sessions on integrating microbial complexity to advance soil carbon predictions! 🦠🌍Invited speakers: Christina Kaiser, Jackie Weissman, Katie Rocci, Will Wieder 🤩. Co-conveners: @ultracricket.bsky.social, @hhollandmoritz.bsky.social, Gianna Marschmann, Jing Tao🙏

November 28, 2024 at 1:29 PM
📢 Add to your #AGU2024 schedule our 3 sessions on integrating microbial complexity to advance soil carbon predictions! 🦠🌍Invited speakers: Christina Kaiser, Jackie Weissman, Katie Rocci, Will Wieder 🤩. Co-conveners: @ultracricket.bsky.social, @hhollandmoritz.bsky.social, Gianna Marschmann, Jing Tao🙏
So honoured to have had Professor Mike Hulme travel from Cambridge under the ❄️ to deliver his talk, "Climate Scientists as Analysts, Advisors, Advocates, or Activists: What’s the Difference and Does It Matter?" @lsce-ipsl.bsky.social
Catch the recording here: www.elsaabs.com/climate-action
Catch the recording here: www.elsaabs.com/climate-action

November 21, 2024 at 6:09 PM
So honoured to have had Professor Mike Hulme travel from Cambridge under the ❄️ to deliver his talk, "Climate Scientists as Analysts, Advisors, Advocates, or Activists: What’s the Difference and Does It Matter?" @lsce-ipsl.bsky.social
Catch the recording here: www.elsaabs.com/climate-action
Catch the recording here: www.elsaabs.com/climate-action
Finally on bluesky!
Talking about integration of microbial evolution into land surface models 🦠🌍
🔗 doi.org/10.1111/gcb....
Talking about integration of microbial evolution into land surface models 🦠🌍
🔗 doi.org/10.1111/gcb....

November 20, 2024 at 10:35 PM
Finally on bluesky!
Talking about integration of microbial evolution into land surface models 🦠🌍
🔗 doi.org/10.1111/gcb....
Talking about integration of microbial evolution into land surface models 🦠🌍
🔗 doi.org/10.1111/gcb....