



Manning Research Group
@manningresearch.bsky.social
Manning Research Group at Syracuse University: theory and computation focused on cells, grains, tissues, glasses, and other out-of-equilibrium disordered matter
Our work confirms that slow tissue movements can generate forces that are significant enough to deform an organ, as the timescale of tissue relaxation is large.
This suggests dynamical forces may be playing a role in many other developmental processes, too. We should look!
13/n
This suggests dynamical forces may be playing a role in many other developmental processes, too. We should look!
13/n
July 17, 2025 at 1:43 PM
Our work confirms that slow tissue movements can generate forces that are significant enough to deform an organ, as the timescale of tissue relaxation is large.
This suggests dynamical forces may be playing a role in many other developmental processes, too. We should look!
13/n
This suggests dynamical forces may be playing a role in many other developmental processes, too. We should look!
13/n
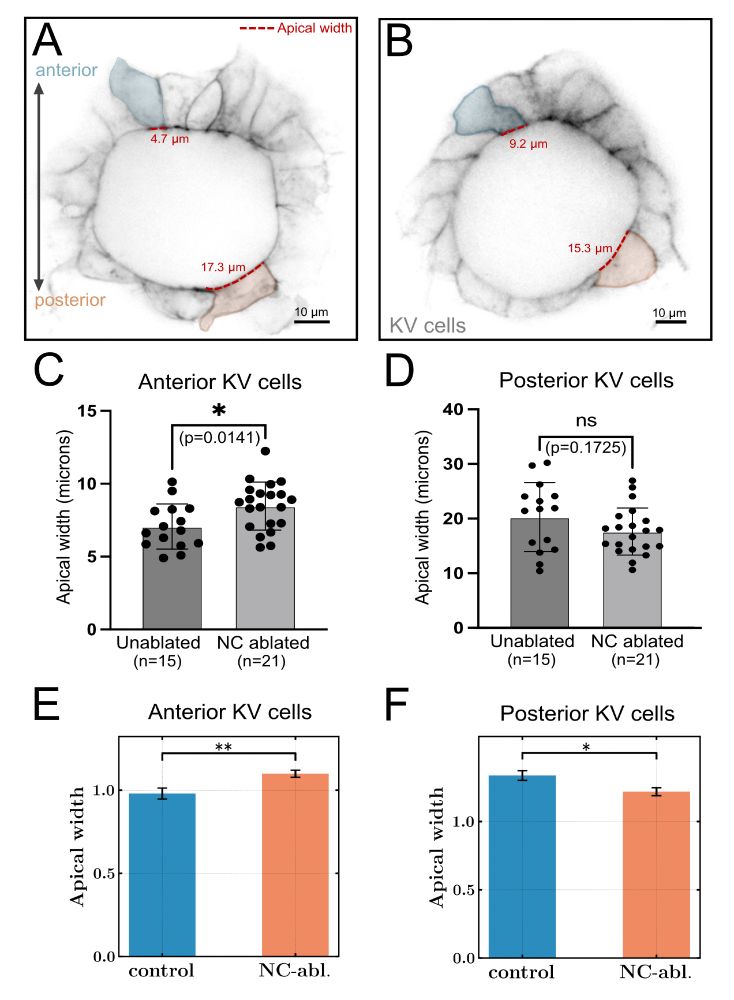
In addition to altering lumen shape changes, are dynamical forces sufficient to change individual cell shapes to drive KV remodeling involved in LR patterning?
Yes, notochord ablation reduces the AP distribution as compared to controls. The 3D vertex model predicts this.
12/n
Yes, notochord ablation reduces the AP distribution as compared to controls. The 3D vertex model predicts this.
12/n
July 17, 2025 at 1:43 PM
In addition to altering lumen shape changes, are dynamical forces sufficient to change individual cell shapes to drive KV remodeling involved in LR patterning?
Yes, notochord ablation reduces the AP distribution as compared to controls. The 3D vertex model predicts this.
12/n
Yes, notochord ablation reduces the AP distribution as compared to controls. The 3D vertex model predicts this.
12/n
These shape changes are generic – they can also be seen in a simpler hydrodynamic model of a membrane surrounded by a highly viscous medium, also with anterior pushing forces and posterior pulling forces.
11/n
11/n
July 17, 2025 at 1:43 PM
These shape changes are generic – they can also be seen in a simpler hydrodynamic model of a membrane surrounded by a highly viscous medium, also with anterior pushing forces and posterior pulling forces.
11/n
11/n
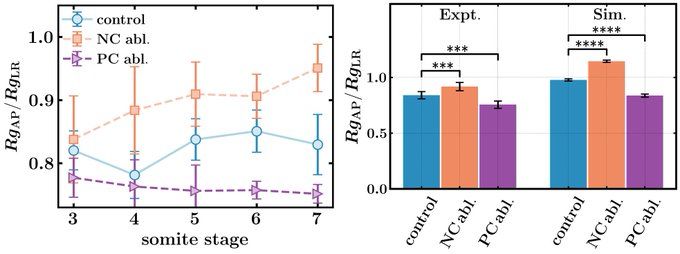
The experiments match the simulation predictions! In notochord ablation experiments, the lumen elongates along the anterior-posterior axis (RgAP/RgLRincreases), while posterior cells ablations extend the lumen along the left-right axis (RgAP/RgLR decreases).
10/n
10/n
July 17, 2025 at 1:43 PM
The experiments match the simulation predictions! In notochord ablation experiments, the lumen elongates along the anterior-posterior axis (RgAP/RgLRincreases), while posterior cells ablations extend the lumen along the left-right axis (RgAP/RgLR decreases).
10/n
10/n
We quantify KV shape by the ratio of the radius of gyration of the lumen along the AP and LR directions (RgAP/RgLR).
In simulations, we mimic notochord (posterior cell) ablations by removing the pushing (pulling) forces, and find that the organ changes it shape significantly.
9/n
In simulations, we mimic notochord (posterior cell) ablations by removing the pushing (pulling) forces, and find that the organ changes it shape significantly.
9/n
July 17, 2025 at 1:43 PM
We quantify KV shape by the ratio of the radius of gyration of the lumen along the AP and LR directions (RgAP/RgLR).
In simulations, we mimic notochord (posterior cell) ablations by removing the pushing (pulling) forces, and find that the organ changes it shape significantly.
9/n
In simulations, we mimic notochord (posterior cell) ablations by removing the pushing (pulling) forces, and find that the organ changes it shape significantly.
9/n
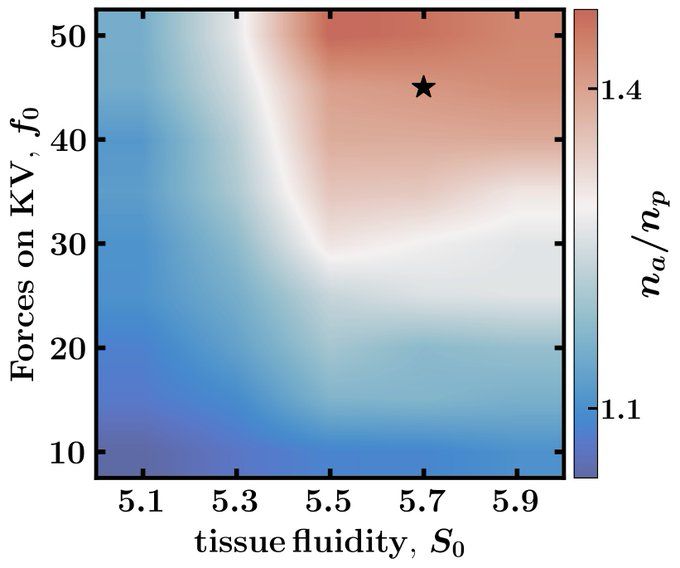
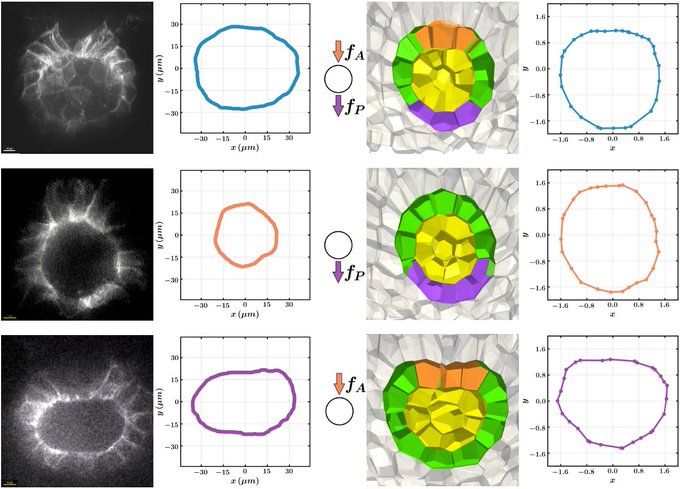
In simulations, we identify a set of model parameters (star in phase diagram) that generate the lumen shape seen in control experiments, where KV is pushed from the anterior by the notochord (orange cells) and pulled by posterior cells (purple cells).
July 17, 2025 at 1:43 PM
In simulations, we identify a set of model parameters (star in phase diagram) that generate the lumen shape seen in control experiments, where KV is pushed from the anterior by the notochord (orange cells) and pulled by posterior cells (purple cells).
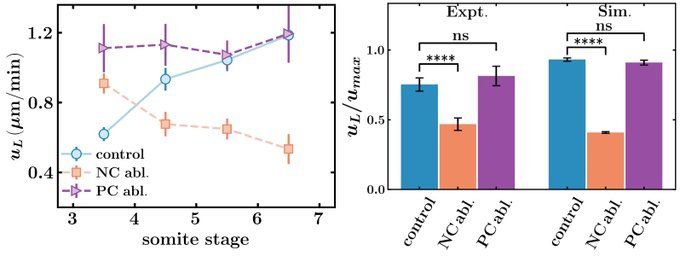
Laser ablation of the notochord reduces the speed of KV in comparison to control experiments. That suggests that we were able to reduce the forces on the KV at the anterior part of KV.
We next ask how these reduced forces affect the shape of the KV lumen and KV cells
7/n
We next ask how these reduced forces affect the shape of the KV lumen and KV cells
7/n
July 17, 2025 at 1:43 PM
Laser ablation of the notochord reduces the speed of KV in comparison to control experiments. That suggests that we were able to reduce the forces on the KV at the anterior part of KV.
We next ask how these reduced forces affect the shape of the KV lumen and KV cells
7/n
We next ask how these reduced forces affect the shape of the KV lumen and KV cells
7/n
To test our hypothesis, we developed 3D models and laser ablation experiments to:
1) quantify how perturbing structures around KV impact its motion
2) measure cell and organ shape in these cases
3) show that observed in vivo shape changes match those predicted from the 3D model
6/n
1) quantify how perturbing structures around KV impact its motion
2) measure cell and organ shape in these cases
3) show that observed in vivo shape changes match those predicted from the 3D model
6/n
July 17, 2025 at 1:43 PM
To test our hypothesis, we developed 3D models and laser ablation experiments to:
1) quantify how perturbing structures around KV impact its motion
2) measure cell and organ shape in these cases
3) show that observed in vivo shape changes match those predicted from the 3D model
6/n
1) quantify how perturbing structures around KV impact its motion
2) measure cell and organ shape in these cases
3) show that observed in vivo shape changes match those predicted from the 3D model
6/n
Our hypothesis: The high-viscosity tailbud tissue exerts drag forces on KV as the organ is pushed through the tissue by convergent extension in the notochord, and pulled through the tissue by posteriorly migrating cells. These forces are sufficient to change KV shape.
5/n
5/n
July 17, 2025 at 1:43 PM
Our hypothesis: The high-viscosity tailbud tissue exerts drag forces on KV as the organ is pushed through the tissue by convergent extension in the notochord, and pulled through the tissue by posteriorly migrating cells. These forces are sufficient to change KV shape.
5/n
5/n
Recent work has demonstrated that many tissues, including zebrafish tailbud, operate close to a fluid-solid transition. Near such transitions, the effective viscosity diverges. In these cases, very slow motions can give rise to large forces.
nature.com/articles/s41...
4/n
nature.com/articles/s41...
4/n
A fluid-to-solid jamming transition underlies vertebrate body axis elongation - Nature
Cell collectives in embryonic tissues undergo a fluid-to-solid jamming transition, similar to those that occur in soft materials such as foams, emulsions and colloidal suspensions, to physically sculp...
nature.com
July 17, 2025 at 1:43 PM
Recent work has demonstrated that many tissues, including zebrafish tailbud, operate close to a fluid-solid transition. Near such transitions, the effective viscosity diverges. In these cases, very slow motions can give rise to large forces.
nature.com/articles/s41...
4/n
nature.com/articles/s41...
4/n
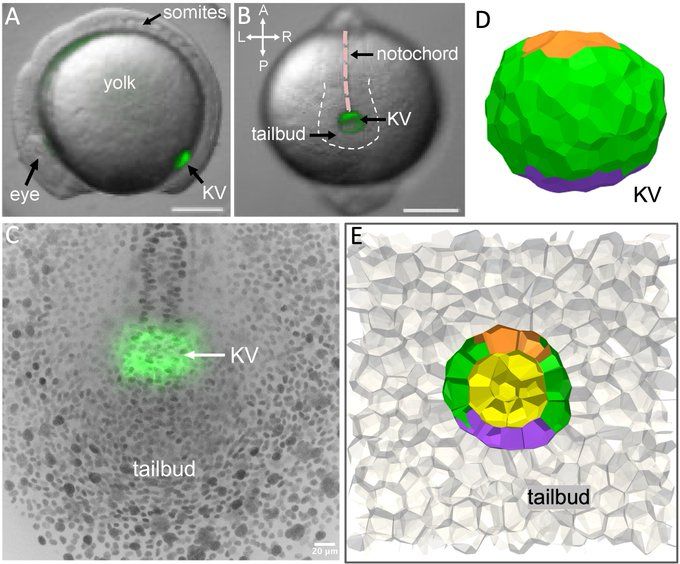
To answer this question, we study Kupffer’s Vesicle (KV). KV is composed of a single layer of epithelial cells surrounding a fluid-filled lumen, located in tailbud of zebrafish, and it moves through surrounding tailbud tissue slowly, at about a micron per minute.
3/n
3/n
July 17, 2025 at 1:43 PM
To answer this question, we study Kupffer’s Vesicle (KV). KV is composed of a single layer of epithelial cells surrounding a fluid-filled lumen, located in tailbud of zebrafish, and it moves through surrounding tailbud tissue slowly, at about a micron per minute.
3/n
3/n
A key question in developmental biology is how organisms robustly control the morphology of tissues and organs. Mechanical forces can help provide such a control mechanism, but because tissue motion is so slow, most studies have assumed local force balance. Is that true?
2/n
2/n
July 17, 2025 at 1:43 PM
A key question in developmental biology is how organisms robustly control the morphology of tissues and organs. Mechanical forces can help provide such a control mechanism, but because tissue motion is so slow, most studies have assumed local force balance. Is that true?
2/n
2/n
This was a fun collaboration spearheaded by @rajphys.bsky.social and Emma Retzlaff, in collaboration with the fabulous Amack Lab and @lovelessradio.bsky.social, in the great environment of @syracuseu.bsky.social
July 17, 2025 at 1:43 PM
This was a fun collaboration spearheaded by @rajphys.bsky.social and Emma Retzlaff, in collaboration with the fabulous Amack Lab and @lovelessradio.bsky.social, in the great environment of @syracuseu.bsky.social
Sure, thanks! www.bigmarker.com/cellpress-so...

Challenges and opportunities for active matter in biology
Join us for an engaging exploration of the dynamic intersection between physics and biology focusing on active matter—non-equilibrium systems of interacting components that consume energy to do work a...
www.bigmarker.com
July 9, 2025 at 5:30 PM
Sure, thanks! www.bigmarker.com/cellpress-so...
What the paper shows is that -- for some key observables -- it doesn't seem to matter which model you choose. The cell shape always predicts the rigidity transition, and the viscoelastic response is the same on either side of the transition.
July 9, 2025 at 5:23 PM
What the paper shows is that -- for some key observables -- it doesn't seem to matter which model you choose. The cell shape always predicts the rigidity transition, and the viscoelastic response is the same on either side of the transition.
But the key point is that we're not sure which of these models is "right" -- some might be right for one cell but not another. So what I mean is that different versions of vertex models are likely "wrong" for certain cell types.
July 9, 2025 at 5:23 PM
But the key point is that we're not sure which of these models is "right" -- some might be right for one cell but not another. So what I mean is that different versions of vertex models are likely "wrong" for certain cell types.
One way of rationalizing the P^2 term is to say some cell types have a contractile actin ring; another way is to say that there's a limited pool in the cell of, e.g. cadherins or myosin.
July 9, 2025 at 5:23 PM
One way of rationalizing the P^2 term is to say some cell types have a contractile actin ring; another way is to say that there's a limited pool in the cell of, e.g. cadherins or myosin.
The linear term is a line tension (2D version of a surface tension) but the quadratic term is more mysterious, and some other versions of vertex models remove it. Other versions of vertex models have a spring-like restoring force on each edge, instead of a restoring force on the perimeter.
July 9, 2025 at 5:23 PM
The linear term is a line tension (2D version of a surface tension) but the quadratic term is more mysterious, and some other versions of vertex models remove it. Other versions of vertex models have a spring-like restoring force on each edge, instead of a restoring force on the perimeter.
We know that cell cortex mechanics is really complex (viscoelastic, with feedback loops, etc.) A standard vertex model makes a specific assumption about that mechanics -- with a term proportional to the perimeter P and a second term proportional to P^2,
July 9, 2025 at 5:23 PM
We know that cell cortex mechanics is really complex (viscoelastic, with feedback loops, etc.) A standard vertex model makes a specific assumption about that mechanics -- with a term proportional to the perimeter P and a second term proportional to P^2,