




Previous experiments observed a transient spike in transcription of a subset of genes during ana/telophase. We now observe that these spiking genes are associated with microcompartments that peak in interactions during ana/telo, suggesting a 3D-genome-based mechanism for this transcriptional spike!

October 17, 2025 at 2:58 PM
Previous experiments observed a transient spike in transcription of a subset of genes during ana/telophase. We now observe that these spiking genes are associated with microcompartments that peak in interactions during ana/telo, suggesting a 3D-genome-based mechanism for this transcriptional spike!
Can PDS5 therefore strengthen CTCF boundaries by limiting NIPBL-cohesin processivity (=speed x residence time)?
Hi-C in cells depleted of PDS5 & now partially of NIPBL recover some of CTCF boundaries (+ also compartments)
Hi-C in cells depleted of PDS5 & now partially of NIPBL recover some of CTCF boundaries (+ also compartments)

September 3, 2025 at 4:20 PM
Can PDS5 therefore strengthen CTCF boundaries by limiting NIPBL-cohesin processivity (=speed x residence time)?
Hi-C in cells depleted of PDS5 & now partially of NIPBL recover some of CTCF boundaries (+ also compartments)
Hi-C in cells depleted of PDS5 & now partially of NIPBL recover some of CTCF boundaries (+ also compartments)
To understand this Iain did 3-color single-molecule expts w/loop-extruding NIPBL-cohesin.
PDS5+NIPBL could colocalize on cohesin leading to NIPBL unbinding! Loops often released but sometimes shrunk & NIPBL returned.
So PDS5 slows loop growth + speeds NIPBL unbinding by facilitated dissociation!
PDS5+NIPBL could colocalize on cohesin leading to NIPBL unbinding! Loops often released but sometimes shrunk & NIPBL returned.
So PDS5 slows loop growth + speeds NIPBL unbinding by facilitated dissociation!

September 3, 2025 at 4:20 PM
To understand this Iain did 3-color single-molecule expts w/loop-extruding NIPBL-cohesin.
PDS5+NIPBL could colocalize on cohesin leading to NIPBL unbinding! Loops often released but sometimes shrunk & NIPBL returned.
So PDS5 slows loop growth + speeds NIPBL unbinding by facilitated dissociation!
PDS5+NIPBL could colocalize on cohesin leading to NIPBL unbinding! Loops often released but sometimes shrunk & NIPBL returned.
So PDS5 slows loop growth + speeds NIPBL unbinding by facilitated dissociation!
We turned to the possibility of PDS5 competing w/NIPBL for cohesin
PDS5 inhibits cohesin-NIPBL ATPase+binding in vitro & suppresses NIPBL occupancy of cohesin in cells(+ PDS5&WAPL dont affect each others occupancy)
iFRAP shows NIPBL chromatin residence time seems to increase a lot w/ PDS5 depletion
PDS5 inhibits cohesin-NIPBL ATPase+binding in vitro & suppresses NIPBL occupancy of cohesin in cells(+ PDS5&WAPL dont affect each others occupancy)
iFRAP shows NIPBL chromatin residence time seems to increase a lot w/ PDS5 depletion

September 3, 2025 at 4:20 PM
We turned to the possibility of PDS5 competing w/NIPBL for cohesin
PDS5 inhibits cohesin-NIPBL ATPase+binding in vitro & suppresses NIPBL occupancy of cohesin in cells(+ PDS5&WAPL dont affect each others occupancy)
iFRAP shows NIPBL chromatin residence time seems to increase a lot w/ PDS5 depletion
PDS5 inhibits cohesin-NIPBL ATPase+binding in vitro & suppresses NIPBL occupancy of cohesin in cells(+ PDS5&WAPL dont affect each others occupancy)
iFRAP shows NIPBL chromatin residence time seems to increase a lot w/ PDS5 depletion
Since PDS5A/B aren't required for cohesin accumulation at CTCF, are they altering TADs etc via SMC3 acetylation? PDS5 depletion reduces acetylation but in separate expts w/ ESCO1/2 depleted we see no acetylation & TADs+CTCF contacts remain. PDS5 not influencing TADs+corner peaks only by acetylation.

September 3, 2025 at 4:20 PM
Since PDS5A/B aren't required for cohesin accumulation at CTCF, are they altering TADs etc via SMC3 acetylation? PDS5 depletion reduces acetylation but in separate expts w/ ESCO1/2 depleted we see no acetylation & TADs+CTCF contacts remain. PDS5 not influencing TADs+corner peaks only by acetylation.
What about CTCF? PDS5A/B isnt needed to recruit CTCF & it aids cohesin accumulation at CTCF. But PDS5-mediated cohesin accumulation isnt merely due to recruitment to CTCF. PDS5 depletion leads to new accumulation of cohesin elsewhere in “cohesin islands” often at sites of convergent transcription.

September 3, 2025 at 4:20 PM
What about CTCF? PDS5A/B isnt needed to recruit CTCF & it aids cohesin accumulation at CTCF. But PDS5-mediated cohesin accumulation isnt merely due to recruitment to CTCF. PDS5 depletion leads to new accumulation of cohesin elsewhere in “cohesin islands” often at sites of convergent transcription.
How does increasing extrusion life hinder or facilitate compartments? Competition w/chromatin conformational dynamics!
Extrusion disrupts contacts. Compartmentalization results from equilibration via polymer relaxation
Fast cohesin turnover->disruptions occur more quickly than they can be relaxed
Extrusion disrupts contacts. Compartmentalization results from equilibration via polymer relaxation
Fast cohesin turnover->disruptions occur more quickly than they can be relaxed


September 3, 2025 at 4:20 PM
How does increasing extrusion life hinder or facilitate compartments? Competition w/chromatin conformational dynamics!
Extrusion disrupts contacts. Compartmentalization results from equilibration via polymer relaxation
Fast cohesin turnover->disruptions occur more quickly than they can be relaxed
Extrusion disrupts contacts. Compartmentalization results from equilibration via polymer relaxation
Fast cohesin turnover->disruptions occur more quickly than they can be relaxed
Why does PDS5A/B depletion erase compartments as well as WAPL depletion despite differences in residence time? One possibility is that PDS5A/B depletion leads to faster average extrusion speed

September 3, 2025 at 4:20 PM
Why does PDS5A/B depletion erase compartments as well as WAPL depletion despite differences in residence time? One possibility is that PDS5A/B depletion leads to faster average extrusion speed
How? In polymer sims increasing cohesin residence time can nonmonotonically alter chromatin compartmentalization. Consistently iFRAP showed residence times increase w/ WAPL & PDS5A/B depletion, dramatically so w/ triple depletion (also suggesting partially distinct PDS5 & WAPL-based cohesin release)


September 3, 2025 at 4:20 PM
How? In polymer sims increasing cohesin residence time can nonmonotonically alter chromatin compartmentalization. Consistently iFRAP showed residence times increase w/ WAPL & PDS5A/B depletion, dramatically so w/ triple depletion (also suggesting partially distinct PDS5 & WAPL-based cohesin release)
As seen previously, PDS5 or WAPL depletion suppress chromatin compartmentalization, but astonishingly, co-depletion of WAPL and PDS5A/B leads to recovery of compartments in near-cis!

September 3, 2025 at 4:20 PM
As seen previously, PDS5 or WAPL depletion suppress chromatin compartmentalization, but astonishingly, co-depletion of WAPL and PDS5A/B leads to recovery of compartments in near-cis!
In auxin-inducible degron cells, Gordana used Hi-C to observe the loss of extrusion-mediated CTCF-CTCF contacts (corner peaks) with PDS5A/B depletion. Interestingly, while WAPL depletion increases corner peaks, co-depleting PDS5 completely suppresses them

September 3, 2025 at 4:20 PM
In auxin-inducible degron cells, Gordana used Hi-C to observe the loss of extrusion-mediated CTCF-CTCF contacts (corner peaks) with PDS5A/B depletion. Interestingly, while WAPL depletion increases corner peaks, co-depleting PDS5 completely suppresses them
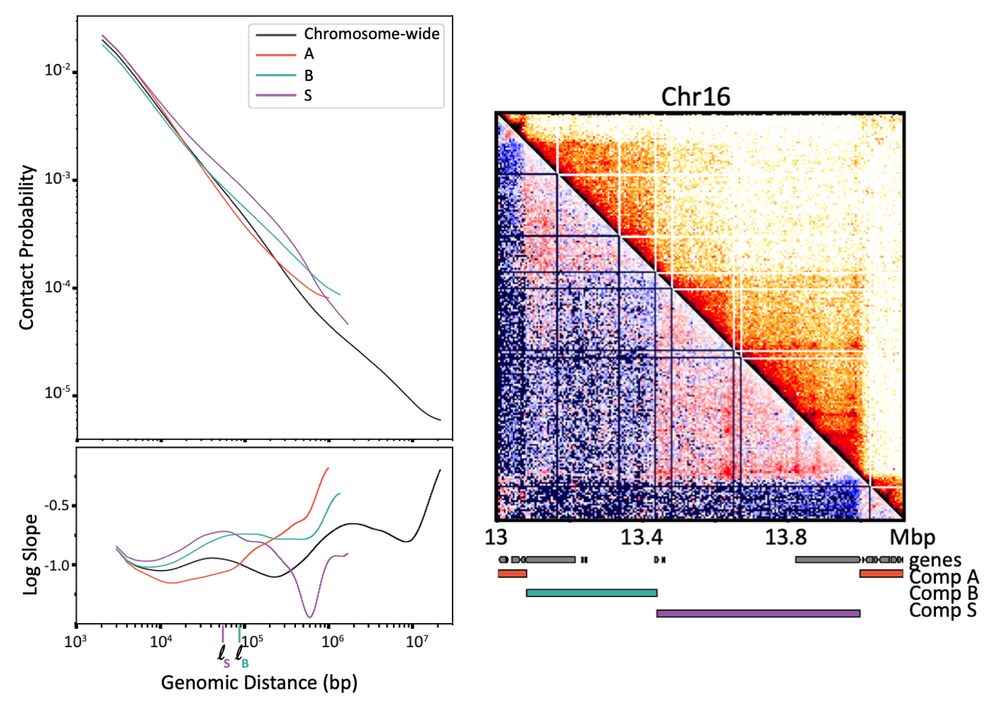
Interestingly, the Hi-C expts display features of loop extrusion - dots of contact enrichment, stripes emanating from S compartment boundaries, and a characteristic extrusion shoulder in the contact frequency curve P(s) that is most notable in S comps
July 8, 2025 at 3:14 PM
Interestingly, the Hi-C expts display features of loop extrusion - dots of contact enrichment, stripes emanating from S compartment boundaries, and a characteristic extrusion shoulder in the contact frequency curve P(s) that is most notable in S comps
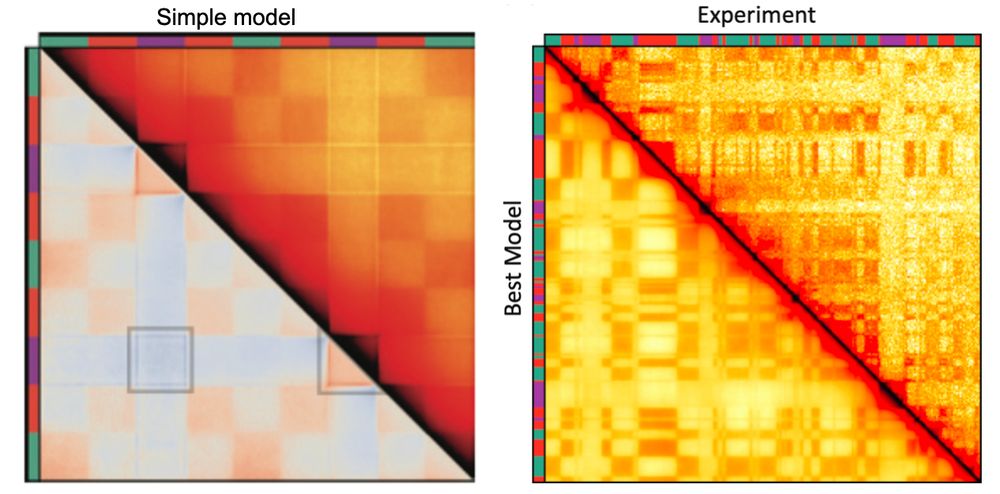
But a cooperative combination of these two mechanisms can generate S compartment patterns! When loop extrusion is targeted to S compartments, we observe local contact enrichment in S, depletion of inter-S contacts, and sequestration from the rest of the chromosome
July 8, 2025 at 3:14 PM
But a cooperative combination of these two mechanisms can generate S compartment patterns! When loop extrusion is targeted to S compartments, we observe local contact enrichment in S, depletion of inter-S contacts, and sequestration from the rest of the chromosome
Ok it’s unusual but is this just microphase separation or loop extrusion? No!
Emily’s simulations show S compartments can't be generated by either of these mechanisms alone! Phase separation makes unwanted inter-S contacts & extrusion can't deplete inter-S contacts w/out erasing all comps
Emily’s simulations show S compartments can't be generated by either of these mechanisms alone! Phase separation makes unwanted inter-S contacts & extrusion can't deplete inter-S contacts w/out erasing all comps

July 8, 2025 at 3:14 PM
Ok it’s unusual but is this just microphase separation or loop extrusion? No!
Emily’s simulations show S compartments can't be generated by either of these mechanisms alone! Phase separation makes unwanted inter-S contacts & extrusion can't deplete inter-S contacts w/out erasing all comps
Emily’s simulations show S compartments can't be generated by either of these mechanisms alone! Phase separation makes unwanted inter-S contacts & extrusion can't deplete inter-S contacts w/out erasing all comps
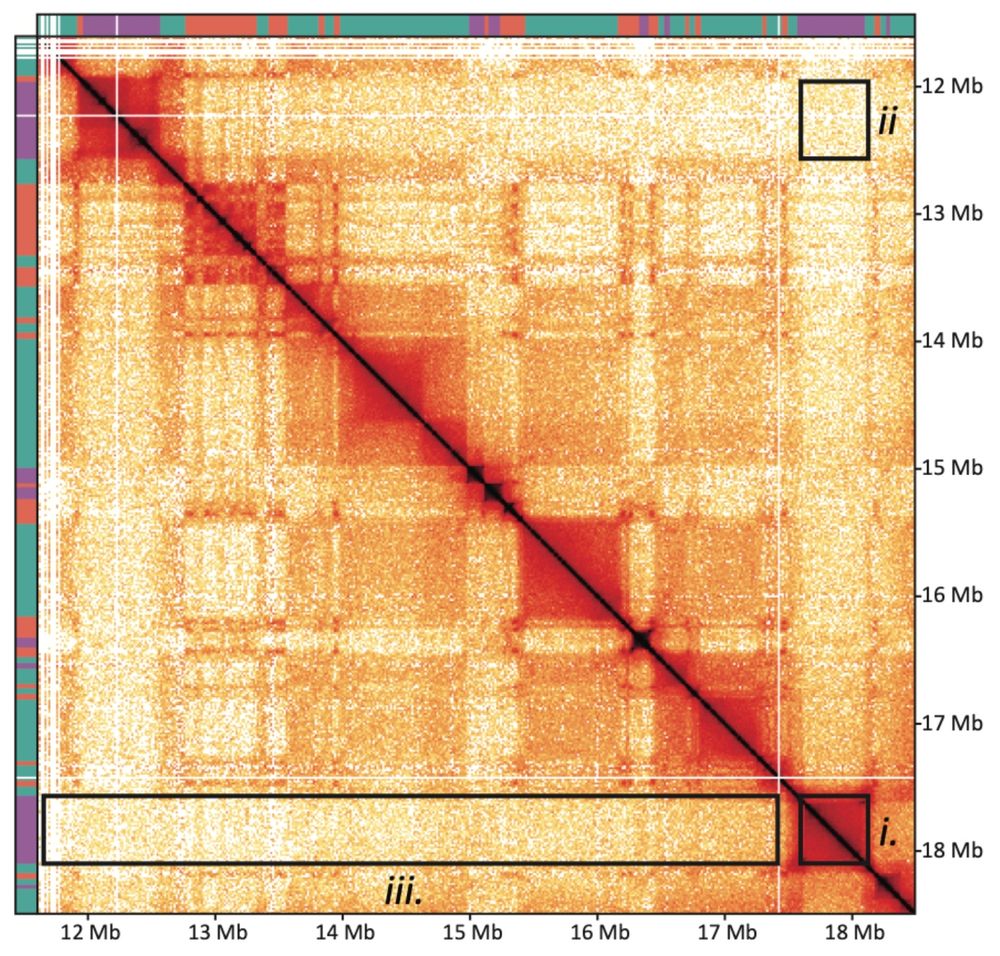
How are S compartments are strange? 3 key features: i) on-diagonal contacts enriched, ii) off-diagonal contact depletion bet/ S comps. (unlike usual compartments, not typical of TADs), iii) smooth depletion of S & all other parts of the chrm (extreme sequestration!)
July 8, 2025 at 3:14 PM
How are S compartments are strange? 3 key features: i) on-diagonal contacts enriched, ii) off-diagonal contact depletion bet/ S comps. (unlike usual compartments, not typical of TADs), iii) smooth depletion of S & all other parts of the chrm (extreme sequestration!)
These physical constraints have implications for force transmission into the nucleus. Upon deformation, tethered & crosslinked peripheral increases contacts with the lamina heterochromatin and reorganizes far more than unconstrained chromatin.

February 17, 2025 at 3:31 PM
These physical constraints have implications for force transmission into the nucleus. Upon deformation, tethered & crosslinked peripheral increases contacts with the lamina heterochromatin and reorganizes far more than unconstrained chromatin.
All changes w/ peripheral heterochromatin tethered to the lamina w/ specific bonds. These tethers stiffen nuclei.
More important, now, increasing phase-separated heterochromatin & crosslinked heterochromatin can increase nuclear stiffness.
Lamina tethers are physical constraints on chromatin!
More important, now, increasing phase-separated heterochromatin & crosslinked heterochromatin can increase nuclear stiffness.
Lamina tethers are physical constraints on chromatin!

February 17, 2025 at 3:31 PM
All changes w/ peripheral heterochromatin tethered to the lamina w/ specific bonds. These tethers stiffen nuclei.
More important, now, increasing phase-separated heterochromatin & crosslinked heterochromatin can increase nuclear stiffness.
Lamina tethers are physical constraints on chromatin!
More important, now, increasing phase-separated heterochromatin & crosslinked heterochromatin can increase nuclear stiffness.
Lamina tethers are physical constraints on chromatin!
But w/ this chromatin model — widely used in 3D genome modeling — increasing heterochromatin *decreases* stiffness rather than increasing it as in expts.
Why? Chromatin condensation reduces polymer osmotic pressure. Stiffening heterochromatin by crosslinking doesnt help either! What’s going on?
Why? Chromatin condensation reduces polymer osmotic pressure. Stiffening heterochromatin by crosslinking doesnt help either! What’s going on?

February 17, 2025 at 3:31 PM
But w/ this chromatin model — widely used in 3D genome modeling — increasing heterochromatin *decreases* stiffness rather than increasing it as in expts.
Why? Chromatin condensation reduces polymer osmotic pressure. Stiffening heterochromatin by crosslinking doesnt help either! What’s going on?
Why? Chromatin condensation reduces polymer osmotic pressure. Stiffening heterochromatin by crosslinking doesnt help either! What’s going on?
We first simulate this model with just heterochromatin (nonspecifically) adsorbed to the lamina and phase-separated from euchromatin.

February 17, 2025 at 3:31 PM
We first simulate this model with just heterochromatin (nonspecifically) adsorbed to the lamina and phase-separated from euchromatin.
Our polymer sim model suggests 3 factors govern microcompartments:
1) affinity interactions
2) loop extrusion
3) chromatin volume density/compaction
Density seems to be particularly important & changes ~2-fold in M-to-G1 transition! But each factor seems to contribute to the observed dynamics
1) affinity interactions
2) loop extrusion
3) chromatin volume density/compaction
Density seems to be particularly important & changes ~2-fold in M-to-G1 transition! But each factor seems to contribute to the observed dynamics

November 8, 2024 at 2:47 PM
Our polymer sim model suggests 3 factors govern microcompartments:
1) affinity interactions
2) loop extrusion
3) chromatin volume density/compaction
Density seems to be particularly important & changes ~2-fold in M-to-G1 transition! But each factor seems to contribute to the observed dynamics
1) affinity interactions
2) loop extrusion
3) chromatin volume density/compaction
Density seems to be particularly important & changes ~2-fold in M-to-G1 transition! But each factor seems to contribute to the observed dynamics
Surprisingly, microcompartment strengths appear to peak during anaphase/telophase and then gradually fade during interphase. This sharply contrasts with observations of larger A/B compartments!

November 8, 2024 at 2:46 PM
Surprisingly, microcompartment strengths appear to peak during anaphase/telophase and then gradually fade during interphase. This sharply contrasts with observations of larger A/B compartments!
Hi-C expts show chromosomes lose well-defined contacts during mitosis—compartments, TADs, etc…but do they really?
Using region capture Micro-C to deeply resolve we find chromosomes form microcompartments of cis-regulatory elements even during mitosis! They appear as a “grid of dots” in contact maps
Using region capture Micro-C to deeply resolve we find chromosomes form microcompartments of cis-regulatory elements even during mitosis! They appear as a “grid of dots” in contact maps

November 8, 2024 at 2:45 PM
Hi-C expts show chromosomes lose well-defined contacts during mitosis—compartments, TADs, etc…but do they really?
Using region capture Micro-C to deeply resolve we find chromosomes form microcompartments of cis-regulatory elements even during mitosis! They appear as a “grid of dots” in contact maps
Using region capture Micro-C to deeply resolve we find chromosomes form microcompartments of cis-regulatory elements even during mitosis! They appear as a “grid of dots” in contact maps