



Greg Keele
@grkeele.bsky.social
The hemoglobin beta locus is driven by a known genetic variant that induces an additional cysteine residue that is present in 5 of the founder strains. This locus regulated glutathione levels and drove a ptmQTL hotspot, highlighting the central role of hemoglobin in RBC metabolism.

March 9, 2025 at 7:34 PM
The hemoglobin beta locus is driven by a known genetic variant that induces an additional cysteine residue that is present in 5 of the founder strains. This locus regulated glutathione levels and drove a ptmQTL hotspot, highlighting the central role of hemoglobin in RBC metabolism.
Likely due to this, we observed a notable lack of cis-genetic regulation compared to other tissues previously studied in these mice. In its place, we observed strong trans pQTL hotspots, including at the hemoglobin alpha and beta. Many align with metabolite and lipid QTL (doi.org/10.1182/bloo...).

March 9, 2025 at 7:34 PM
Likely due to this, we observed a notable lack of cis-genetic regulation compared to other tissues previously studied in these mice. In its place, we observed strong trans pQTL hotspots, including at the hemoglobin alpha and beta. Many align with metabolite and lipid QTL (doi.org/10.1182/bloo...).
We profiled the red blood cell (RBC) proteome (including PTMs) and the impacts of RBC storage in a genetically diverse mouse population. The RBC is a fascinating and unique cell system, notably lacking nuclei and the ability to respond to stress via de novo protein synthesis.

March 9, 2025 at 7:34 PM
We profiled the red blood cell (RBC) proteome (including PTMs) and the impacts of RBC storage in a genetically diverse mouse population. The RBC is a fascinating and unique cell system, notably lacking nuclei and the ability to respond to stress via de novo protein synthesis.
Please check out heavily revised version of my manuscript comparing genetically diverse mouse populations (CC, CC-RIX, DO). I greatly expanded the simulations and now provide power curves!
x.com/grkeele/status…
x.com/grkeele/status…

March 8, 2025 at 1:59 AM
Please check out heavily revised version of my manuscript comparing genetically diverse mouse populations (CC, CC-RIX, DO). I greatly expanded the simulations and now provide power curves!
x.com/grkeele/status…
x.com/grkeele/status…
Other interesting findings include how CC-RIX samples with replicates can better estimate additive heritability in the presence of F1-specific genetic effects. The CC can't tease these components apart, and when trying to estimate just the additive, returns the sum.

March 8, 2025 at 1:58 AM
Other interesting findings include how CC-RIX samples with replicates can better estimate additive heritability in the presence of F1-specific genetic effects. The CC can't tease these components apart, and when trying to estimate just the additive, returns the sum.
In contrast, I expected the DO to be better at QTL mapping, particularly for small effect loci in polygenic traits. The extent of this was also surprising, with 174 DO mice being better power to detect a 10% QTL in a trait that is 90% heritable than 500 CC or CC-RIX mice.

March 8, 2025 at 1:58 AM
In contrast, I expected the DO to be better at QTL mapping, particularly for small effect loci in polygenic traits. The extent of this was also surprising, with 174 DO mice being better power to detect a 10% QTL in a trait that is 90% heritable than 500 CC or CC-RIX mice.
My hunch was that experiments that included replicates (CC or CC-RIX) would more efficiently estimate heritability. The extent of this surprised me. For example, five replicates per 10 CC strains (50 mice total) had greater precision than 500 DO mice in some cases!

March 8, 2025 at 1:58 AM
My hunch was that experiments that included replicates (CC or CC-RIX) would more efficiently estimate heritability. The extent of this surprised me. For example, five replicates per 10 CC strains (50 mice total) had greater precision than 500 DO mice in some cases!
Interested in doing an genetic experiment with a cutting-edge mouse multiparental population (MPP), but don't know which best suits your needs (e.g., inbred vs outbred)?
Using simulations from real genetic data, I sought to provide some answers and guidelines.
x.com/biorxivpreprin…
Using simulations from real genetic data, I sought to provide some answers and guidelines.
x.com/biorxivpreprin…

March 8, 2025 at 1:58 AM
Interested in doing an genetic experiment with a cutting-edge mouse multiparental population (MPP), but don't know which best suits your needs (e.g., inbred vs outbred)?
Using simulations from real genetic data, I sought to provide some answers and guidelines.
x.com/biorxivpreprin…
Using simulations from real genetic data, I sought to provide some answers and guidelines.
x.com/biorxivpreprin…
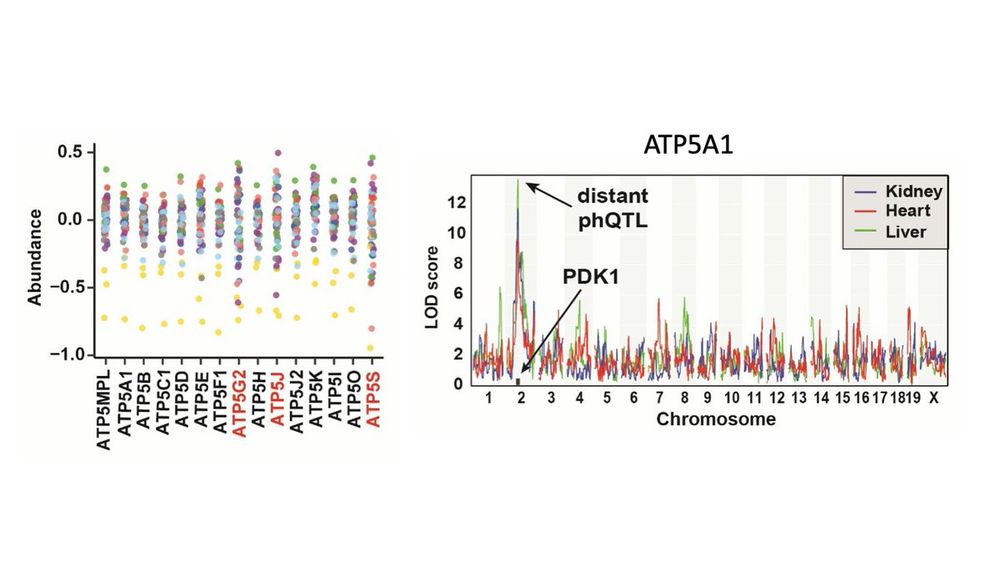
Other cool stories abound. Abundance of the ATP synthase in the heart appears to be stoichiometrically regulated through a low AJ allele at ATP5H. Independent of its protein abundance, phosphorylation of ATP5A1 is regulated by Pdk1.
March 8, 2025 at 1:58 AM
Other cool stories abound. Abundance of the ATP synthase in the heart appears to be stoichiometrically regulated through a low AJ allele at ATP5H. Independent of its protein abundance, phosphorylation of ATP5A1 is regulated by Pdk1.
The phos regulation through Pdk1 is particularly interesting because it is primarily driven by a low NZO allele. The NZO mouse is a very distinct polygenic model for obesity and diabetes, suggesting that unique regulation of phosphorylation could contribute to its phenotype.

March 8, 2025 at 1:58 AM
The phos regulation through Pdk1 is particularly interesting because it is primarily driven by a low NZO allele. The NZO mouse is a very distinct polygenic model for obesity and diabetes, suggesting that unique regulation of phosphorylation could contribute to its phenotype.
We looked for genetic effects independent of parent protein by using a regression adjustment. This allowed us to identify distant phQTL that were mediated by plausible catalysts, such as kinases. For example, Pdk1 appears to phosphorylate 7 proteins across the tissues.

March 8, 2025 at 1:58 AM
We looked for genetic effects independent of parent protein by using a regression adjustment. This allowed us to identify distant phQTL that were mediated by plausible catalysts, such as kinases. For example, Pdk1 appears to phosphorylate 7 proteins across the tissues.
The abundance of a phosphopeptide was often determined by the abundance of its protein (which we refer to as the parent protein). Using a form of mediation analysis, we see that many phQTL essentially reflect an underlying pQTL, particularly when local.

March 8, 2025 at 1:58 AM
The abundance of a phosphopeptide was often determined by the abundance of its protein (which we refer to as the parent protein). Using a form of mediation analysis, we see that many phQTL essentially reflect an underlying pQTL, particularly when local.
This project builds from our previous work where we compared genetic effects on liver proteins among genetically diverse mouse populations.
Here, we integrate expression, proteins, and phosphopeptides from three tissues of 58 inbred CC mouse strains.
doi.org/10.1016/j.xgen…
Here, we integrate expression, proteins, and phosphopeptides from three tissues of 58 inbred CC mouse strains.
doi.org/10.1016/j.xgen…

March 8, 2025 at 1:58 AM
This project builds from our previous work where we compared genetic effects on liver proteins among genetically diverse mouse populations.
Here, we integrate expression, proteins, and phosphopeptides from three tissues of 58 inbred CC mouse strains.
doi.org/10.1016/j.xgen…
Here, we integrate expression, proteins, and phosphopeptides from three tissues of 58 inbred CC mouse strains.
doi.org/10.1016/j.xgen…
The effects of age and sex on proteins complexes can be, well, complex. We see changes in overall abundance due to age and sex, as well as changes to how tightly correlated complex members are. These differences can also be tissue-specific, as in the following ribosome complex.

March 8, 2025 at 1:57 AM
The effects of age and sex on proteins complexes can be, well, complex. We see changes in overall abundance due to age and sex, as well as changes to how tightly correlated complex members are. These differences can also be tissue-specific, as in the following ribosome complex.
Tissues also exhibit unique aging changes to functionally related proteins. In the spleen, for example, proteins increase with age that are related to the ER and protein trafficking. This also highlights co-regulatory signatures we see for protein complexes.

March 8, 2025 at 1:57 AM
Tissues also exhibit unique aging changes to functionally related proteins. In the spleen, for example, proteins increase with age that are related to the ER and protein trafficking. This also highlights co-regulatory signatures we see for protein complexes.
Across tissues, we see aging changes in immune protein levels. Immunoglobulins notably tend to increase with age. Even in tissues where the differences don't meet statistical significance, the direction of effects are consistent. We also see matching signal in immunoproteasomes.

March 8, 2025 at 1:57 AM
Across tissues, we see aging changes in immune protein levels. Immunoglobulins notably tend to increase with age. Even in tissues where the differences don't meet statistical significance, the direction of effects are consistent. We also see matching signal in immunoproteasomes.
The majority of age differences for proteins were not observed in their transcripts. This is in contrast to sex differences, which are often consistent.
We compared to transcript data from a separate study of a sister strain of B6. This matches our previous findings in DO mice.
We compared to transcript data from a separate study of a sister strain of B6. This matches our previous findings in DO mice.

March 8, 2025 at 1:57 AM
The majority of age differences for proteins were not observed in their transcripts. This is in contrast to sex differences, which are often consistent.
We compared to transcript data from a separate study of a sister strain of B6. This matches our previous findings in DO mice.
We compared to transcript data from a separate study of a sister strain of B6. This matches our previous findings in DO mice.
We tested for differences in individual proteins' abundance due to age, sex, and age-by-sex. Tissues like kidney and liver had many proteins with sex differences, consistent with previous studies. Notably, many tissues had many proteins (approaching 500) with age differences.

March 8, 2025 at 1:57 AM
We tested for differences in individual proteins' abundance due to age, sex, and age-by-sex. Tissues like kidney and liver had many proteins with sex differences, consistent with previous studies. Notably, many tissues had many proteins (approaching 500) with age differences.
We used multiplexed mass-spec proteomics to quantify protein abundance across 10 tissues from 20 C57BL/6J mice, representing a balanced factorial design in terms of sex and age (8 and 18 months) within a tissue. Age groups are roughly analogous to young adult and later midlife.

March 8, 2025 at 1:57 AM
We used multiplexed mass-spec proteomics to quantify protein abundance across 10 tissues from 20 C57BL/6J mice, representing a balanced factorial design in terms of sex and age (8 and 18 months) within a tissue. Age groups are roughly analogous to young adult and later midlife.
A key feature is flexibility in inputs for X. In genetically diverse mice, we mapped a distal pQTL for the gene Snx4, which is likely mediated by the related gene Snx7. Inputing X as haplotype data rather than single variants finds strong support for complete mediation.

March 8, 2025 at 1:57 AM
A key feature is flexibility in inputs for X. In genetically diverse mice, we mapped a distal pQTL for the gene Snx4, which is likely mediated by the related gene Snx7. Inputing X as haplotype data rather than single variants finds strong support for complete mediation.
Consider a QTL, where genetic variation at a locus (X) controls phenotypes M and Y. Ideally, mediation analysis can elucidate the relationship between M and Y. We show how different causal relationships between simulated X, M, and Y appear as QTL and our mediation results.

March 8, 2025 at 1:57 AM
Consider a QTL, where genetic variation at a locus (X) controls phenotypes M and Y. Ideally, mediation analysis can elucidate the relationship between M and Y. We show how different causal relationships between simulated X, M, and Y appear as QTL and our mediation results.
We leveraged the replicate CC observations to identify unique CC strain protein dynamics, some due to known genetic variation in a CC strain (such as C3 in CC026). We also identified functionally-enriched sets of proteins for CC strains, highlighting strain-specific biology.

March 8, 2025 at 1:56 AM
We leveraged the replicate CC observations to identify unique CC strain protein dynamics, some due to known genetic variation in a CC strain (such as C3 in CC026). We also identified functionally-enriched sets of proteins for CC strains, highlighting strain-specific biology.
Differences due occur between the CC and DO, notably at the level of emergent phenotypes, like protein complexes. For the exosome complex in CC, genetic variation at one member regulates the entire complex. This is absent in the DO, where the homozygous genotype is not observed.

March 8, 2025 at 1:56 AM
Differences due occur between the CC and DO, notably at the level of emergent phenotypes, like protein complexes. For the exosome complex in CC, genetic variation at one member regulates the entire complex. This is absent in the DO, where the homozygous genotype is not observed.
We quantified abundance for >6000 proteins from the livers of male/female pairs of 58 Collaborative Cross (CC) mouse strains, using multiplexed mass-spec. We then characterized the effects of sex and genetics on proteins and protein complexes.

March 8, 2025 at 1:56 AM
We quantified abundance for >6000 proteins from the livers of male/female pairs of 58 Collaborative Cross (CC) mouse strains, using multiplexed mass-spec. We then characterized the effects of sex and genetics on proteins and protein complexes.
Fun collaboration with @ahepperla and the @DrBrianStrahl lab. Studied rapid H3K36me dynamics genome-wide in yeast using optogenetic switch. We also used Bayesian modeling to detect genes with consistent patterns, accounting for non-linear data and replicate observations.

March 8, 2025 at 1:56 AM
Fun collaboration with @ahepperla and the @DrBrianStrahl lab. Studied rapid H3K36me dynamics genome-wide in yeast using optogenetic switch. We also used Bayesian modeling to detect genes with consistent patterns, accounting for non-linear data and replicate observations.