



Anton Goloborodko
@golobor.bsky.social
A bio-physicist turned phys-biologist,
building models and software in genome biology.
3D genome structure in mitosis | DNA repair | meiosis.
A group leader at @IMBA_Vienna.
Dad x2.
building models and software in genome biology.
3D genome structure in mitosis | DNA repair | meiosis.
A group leader at @IMBA_Vienna.
Dad x2.
he's left-handed!

September 30, 2025 at 1:26 PM
he's left-handed!
something like this?..

July 15, 2025 at 3:03 PM
something like this?..
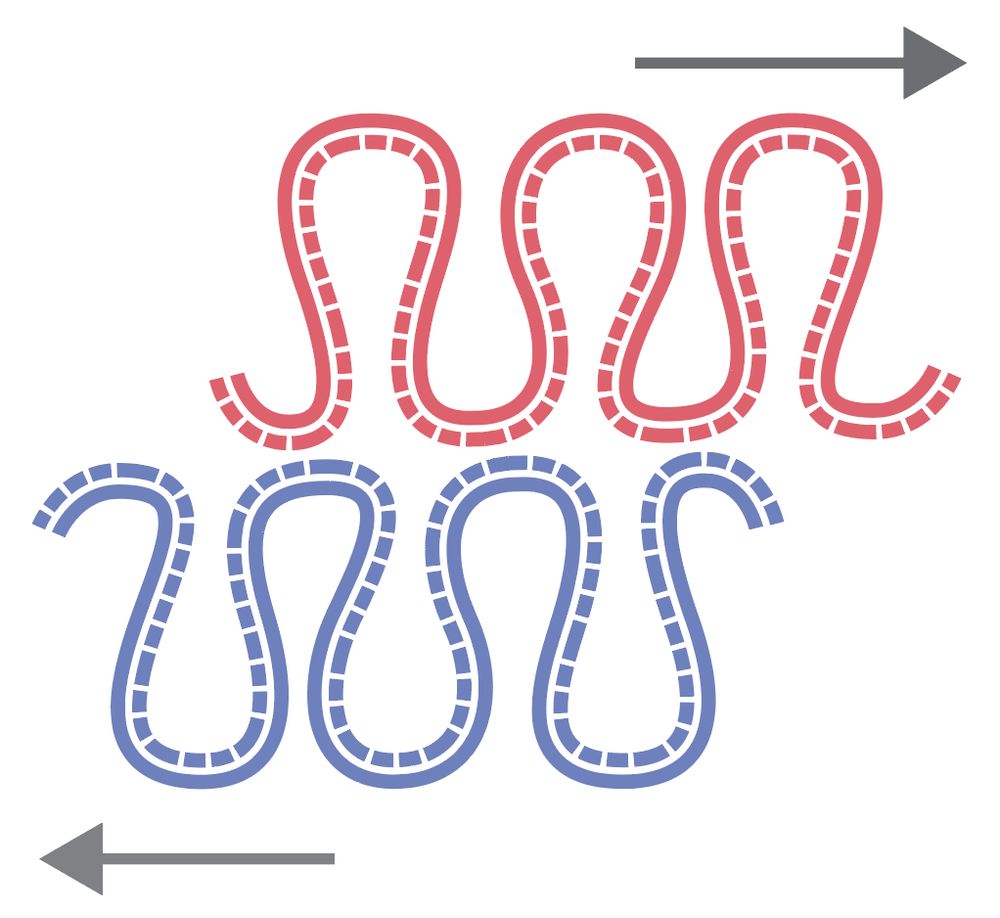
Model 2: cohesins are semi-anchored - fixed to one sister, free on the other. Replication anchors cohesins to leading strands and creates pairs anchored to opposite chromatids. These pairs slide apart but cannot cross, generating the same directional bias without active fork pushing. 12/

July 15, 2025 at 8:11 AM
Model 2: cohesins are semi-anchored - fixed to one sister, free on the other. Replication anchors cohesins to leading strands and creates pairs anchored to opposite chromatids. These pairs slide apart but cannot cross, generating the same directional bias without active fork pushing. 12/
We propose that this asymmetry is “injected” into cohesin alignment during cohesion establishment by asymmetric replication forks.
Model 1: cohesins are pushed/dragged by forks along the leading strand while staying put on the lagging strand, generating the offset. 11/
Model 1: cohesins are pushed/dragged by forks along the leading strand while staying put on the lagging strand, generating the offset. 11/

July 15, 2025 at 8:11 AM
We propose that this asymmetry is “injected” into cohesin alignment during cohesion establishment by asymmetric replication forks.
Model 1: cohesins are pushed/dragged by forks along the leading strand while staying put on the lagging strand, generating the offset. 11/
Model 1: cohesins are pushed/dragged by forks along the leading strand while staying put on the lagging strand, generating the offset. 11/
Our polymer simulations show that the ~2x directional bias in Hi-C data could arise when cohesive cohesins bridge sites are mis-registered by ~100 kb, predominantly in the “1<2” direction. 10/

July 15, 2025 at 8:11 AM
Our polymer simulations show that the ~2x directional bias in Hi-C data could arise when cohesive cohesins bridge sites are mis-registered by ~100 kb, predominantly in the “1<2” direction. 10/
What maintains the shift? Sister chromatids are held together by “cohesive” cohesins, while another “extrusive” cohesin pool forms loops in cis. We killed cohesin loop extrusion by removing NIPBL, but the shift persisted. Depleting cohesive cohesins, however, reduced/erased the asymmetry/shift! 9/

July 15, 2025 at 8:11 AM
What maintains the shift? Sister chromatids are held together by “cohesive” cohesins, while another “extrusive” cohesin pool forms loops in cis. We killed cohesin loop extrusion by removing NIPBL, but the shift persisted. Depleting cohesive cohesins, however, reduced/erased the asymmetry/shift! 9/
Across three regions, probes in the “1 < 2” arrangement are 100–200 nm closer than those in the opposite, “2 < 1” arrangement. Imaging thus confirms what we see in Hi-C: sister chromatids are physically shifted in the 5′→3′ direction of the inherited strands. 8/

July 15, 2025 at 8:11 AM
Across three regions, probes in the “1 < 2” arrangement are 100–200 nm closer than those in the opposite, “2 < 1” arrangement. Imaging thus confirms what we see in Hi-C: sister chromatids are physically shifted in the 5′→3′ direction of the inherited strands. 8/
Thomas Steinacker from the Gerlich lab verified this using optical microscopy and RASER-FISH. He hybridized strand-specific probes, measured distances between sister loci, and visualized the shift. 7/

July 15, 2025 at 8:11 AM
Thomas Steinacker from the Gerlich lab verified this using optical microscopy and RASER-FISH. He hybridized strand-specific probes, measured distances between sister loci, and visualized the shift. 7/
Trans-sister contacts show a ~2-fold enrichment of the “1 < 2” orientation from 100 kb to 3 Mb, peaking near 1 Mb and present in >97 % of the genome. 6/

July 15, 2025 at 8:11 AM
Trans-sister contacts show a ~2-fold enrichment of the “1 < 2” orientation from 100 kb to 3 Mb, peaking near 1 Mb and present in >97 % of the genome. 6/
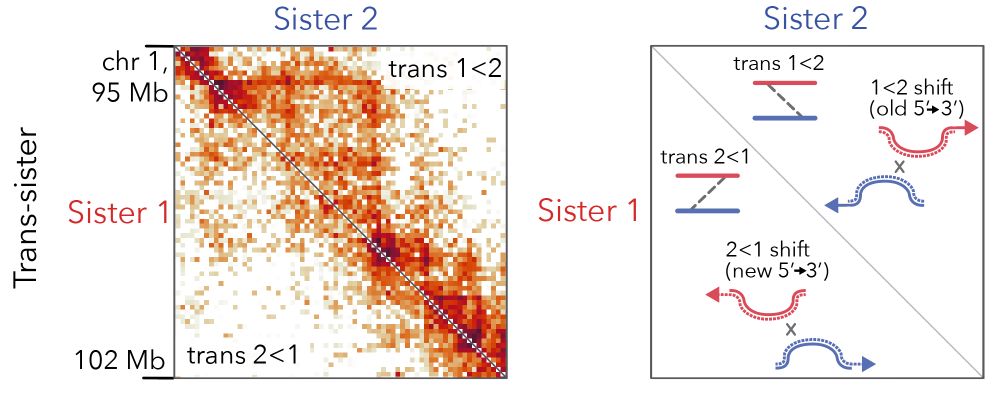
The surprise came from trans-sister maps: contacts above the diagonal - lower-coordinate loci on Sister 1 vs higher-coordinate loci on Sister 2 - outnumber the reverse. An asymmetric/biased matrix implies a genome-wide register shift between sisters in the 5->3 direction of inherited strands!! 5/
July 15, 2025 at 8:11 AM
The surprise came from trans-sister maps: contacts above the diagonal - lower-coordinate loci on Sister 1 vs higher-coordinate loci on Sister 2 - outnumber the reverse. An asymmetric/biased matrix implies a genome-wide register shift between sisters in the 5->3 direction of inherited strands!! 5/
The cis contact maps for the two sisters looked very similar. Contact patterns, P(s) curves and CTCF-anchored loops - all match between the sisters. Within our resolution, replicated sisters seem to have the same loop architecture. 4/

July 15, 2025 at 8:11 AM
The cis contact maps for the two sisters looked very similar. Contact patterns, P(s) curves and CTCF-anchored loops - all match between the sisters. Within our resolution, replicated sisters seem to have the same loop architecture. 4/
We figured one can answer this Q using sister-chromatid-sensitive Hi-C: label new DNA strands with 4sT, call strand-specific mutations, and assign reads to sisters, depending on which strand got labelled. This lets us split Hi-C contacts between Sisters 1 and 2 (inheriting the + and - strands). 3/

July 15, 2025 at 8:11 AM
We figured one can answer this Q using sister-chromatid-sensitive Hi-C: label new DNA strands with 4sT, call strand-specific mutations, and assign reads to sisters, depending on which strand got labelled. This lets us split Hi-C contacts between Sisters 1 and 2 (inheriting the + and - strands). 3/
DNA replication is intrinsically asymmetric: each sister keeps a different parental strand, while new strands are synthesized by asymmetric forks with different leading-/lagging-strand machineries. But does this molecular asymmetry affect large-scale chromosome architecture? 2/

July 15, 2025 at 8:11 AM
DNA replication is intrinsically asymmetric: each sister keeps a different parental strand, while new strands are synthesized by asymmetric forks with different leading-/lagging-strand machineries. But does this molecular asymmetry affect large-scale chromosome architecture? 2/
We found a new asymmetry in the large-scale chromosome structure: sister chromatids are systematically shifted by hundreds of kb in the 5′→3′ direction of their inherited strands! The work was led by Flavia Corsi, in close collaboration with the Daniel Gerlich lab.
www.biorxiv.org/content/10.1...
1/
www.biorxiv.org/content/10.1...
1/
July 15, 2025 at 8:11 AM
We found a new asymmetry in the large-scale chromosome structure: sister chromatids are systematically shifted by hundreds of kb in the 5′→3′ direction of their inherited strands! The work was led by Flavia Corsi, in close collaboration with the Daniel Gerlich lab.
www.biorxiv.org/content/10.1...
1/
www.biorxiv.org/content/10.1...
1/
16/ (iv) in cohesin-depleted mutants, condensins I & II act additively and in parallel, forming nested loops with the same parameters as in chromosomes, built by single condensins


April 11, 2025 at 7:53 AM
16/ (iv) in cohesin-depleted mutants, condensins I & II act additively and in parallel, forming nested loops with the same parameters as in chromosomes, built by single condensins
14/ These models taught us that:
(i) condensin II scaffold is discontinuous with gaps between loops
(ii) strong second diagonals in population Hi-C can be explained by weak, irregular condensin II-mediated spiraling consistent with microscopy data
(i) condensin II scaffold is discontinuous with gaps between loops
(ii) strong second diagonals in population Hi-C can be explained by weak, irregular condensin II-mediated spiraling consistent with microscopy data

April 11, 2025 at 7:53 AM
14/ These models taught us that:
(i) condensin II scaffold is discontinuous with gaps between loops
(ii) strong second diagonals in population Hi-C can be explained by weak, irregular condensin II-mediated spiraling consistent with microscopy data
(i) condensin II scaffold is discontinuous with gaps between loops
(ii) strong second diagonals in population Hi-C can be explained by weak, irregular condensin II-mediated spiraling consistent with microscopy data
13/ These detailed views of mitotic chromosome structure allowed us to design & fit next-gen polymer models for chromosomes formed by condensins I, II & both together. The new models accurately reproduce Hi-C contact frequencies from kb to tens of Mb.



April 11, 2025 at 7:53 AM
13/ These detailed views of mitotic chromosome structure allowed us to design & fit next-gen polymer models for chromosomes formed by condensins I, II & both together. The new models accurately reproduce Hi-C contact frequencies from kb to tens of Mb.
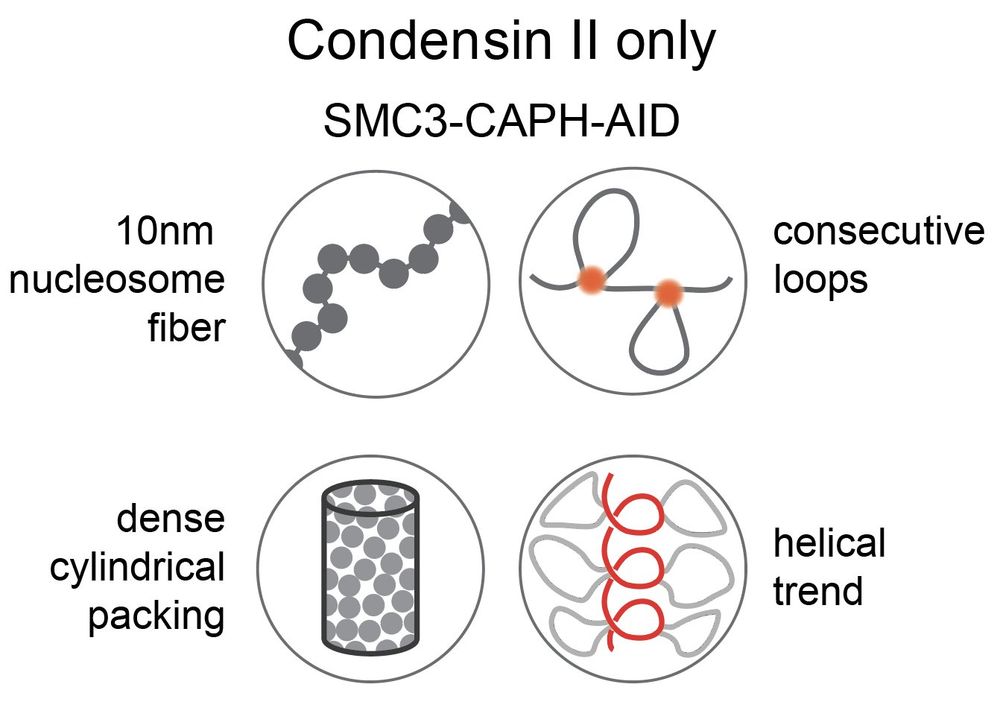
12/ Condensin II-only chromosomes had striking Hi-C maps with a robust third diagonal & fainter 4th diagonal. The second diagonal shifted to ~16 Mb (vs. ~8 Mb for condensin I+II), revealing more DNA per helical gyre.

April 11, 2025 at 7:53 AM
12/ Condensin II-only chromosomes had striking Hi-C maps with a robust third diagonal & fainter 4th diagonal. The second diagonal shifted to ~16 Mb (vs. ~8 Mb for condensin I+II), revealing more DNA per helical gyre.
11/ Lack of cohesin exaggerated differences in Hi-C data for condensin I-only & condensin II-only chromosomes. Condensin I-only chromosomes lacked the second diagonal, indicating no helical organization.

April 11, 2025 at 7:53 AM
11/ Lack of cohesin exaggerated differences in Hi-C data for condensin I-only & condensin II-only chromosomes. Condensin I-only chromosomes lacked the second diagonal, indicating no helical organization.
10/ These phenotypes were more exaggerated than in previously studied single condensin depletions in the presence of intact cohesins. SBF-SEM allowed 3D reconstruction & volume measurement of chromosomes at extremely high resolution.

April 11, 2025 at 7:53 AM
10/ These phenotypes were more exaggerated than in previously studied single condensin depletions in the presence of intact cohesins. SBF-SEM allowed 3D reconstruction & volume measurement of chromosomes at extremely high resolution.
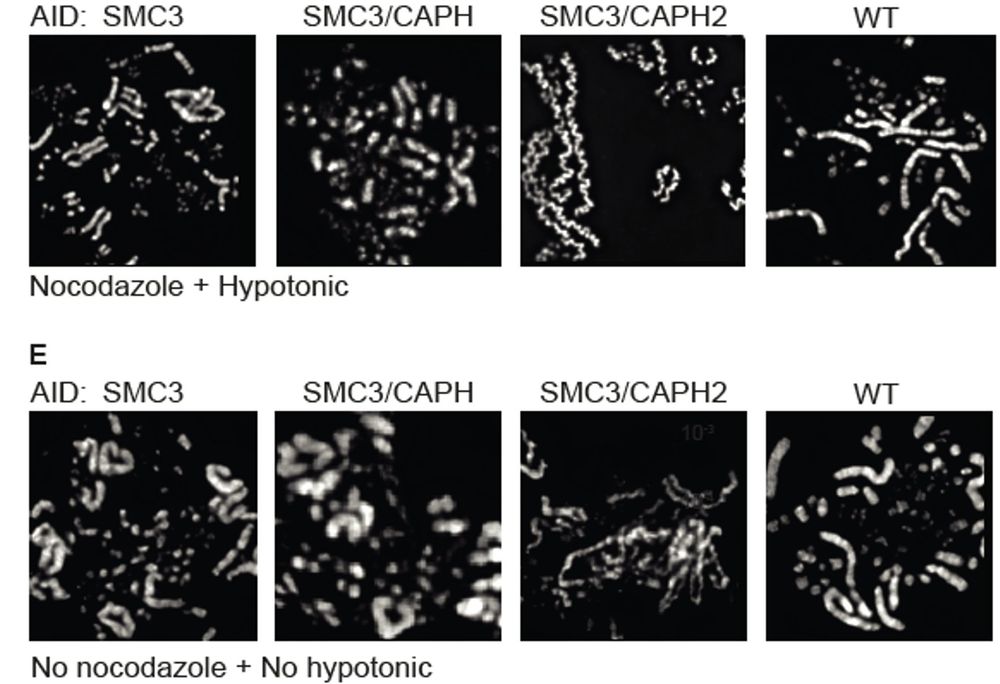
9/ Microscopy revealed short, thick chromatids in SMC3-CAPH (only condensin II active), while SMC3-CAPH2 (only condensin I) had long, thin, disorganized chromosomes.
April 11, 2025 at 7:53 AM
9/ Microscopy revealed short, thick chromatids in SMC3-CAPH (only condensin II active), while SMC3-CAPH2 (only condensin I) had long, thin, disorganized chromosomes.
7/ These "double synchronization" experiments showed cohesive, not extrusive, cohesins affect mitotic chromosome shape by limiting each sister's ability to form an independent helical condensin loop array.

April 11, 2025 at 7:53 AM
7/ These "double synchronization" experiments showed cohesive, not extrusive, cohesins affect mitotic chromosome shape by limiting each sister's ability to form an independent helical condensin loop array.
6/ To test if "extrusive" or "cohesive" cohesins affect wild-type chromosome conformation, we temporarily depleted all cohesins in G2 and then brought them back. Since cohesion is established in S-phase, only extrusive cohesins return in G2.

April 11, 2025 at 7:53 AM
6/ To test if "extrusive" or "cohesive" cohesins affect wild-type chromosome conformation, we temporarily depleted all cohesins in G2 and then brought them back. Since cohesion is established in S-phase, only extrusive cohesins return in G2.
5/ Cohesin depletion alters chromosome morphology: separated sisters become shorter & wider, and Hi-C maps show a brighter "second diagonal", likely due to more pronounced internal helicity w/o cohesin.


April 11, 2025 at 7:53 AM
5/ Cohesin depletion alters chromosome morphology: separated sisters become shorter & wider, and Hi-C maps show a brighter "second diagonal", likely due to more pronounced internal helicity w/o cohesin.
4/ In WT prometaphase, cohesins accumulate between sister chromatids. Models suggest those are cohesive cohesins holding sister chromatids together and that extrusive condensins must bypass these cohesins, allowing chromatid individualization while preserving sister cohesion.


April 11, 2025 at 7:53 AM
4/ In WT prometaphase, cohesins accumulate between sister chromatids. Models suggest those are cohesive cohesins holding sister chromatids together and that extrusive condensins must bypass these cohesins, allowing chromatid individualization while preserving sister cohesion.